Faculty of Pharmacy, Uttar Pradesh University of Medical Sciences, Saifai, Etawah 206130, UP, India.
Neurological problems known as autism have an impact on communication and social interaction. recurring actions, social interactions, and spoken and nonverbal communication. Further to increased anxiety and cognitive deficits is the type of hereditary intellectual disability that is most prevalent. No medicine exist that are effective in treating the main symptoms of autism spectrum disorder. Basically, the factors causing their being are currently unsolved. There are limited animal models of autism diseases that replicate one or a few of the main symptoms of the problem (Social behavior infractions and relative communication conditions). elevated plus maze, open-field, self-grooming, and new item identification behavioral testing. It was established that neurotrophin-related methods and a number of autism-like symptoms are connected. New genetic animal models of the behavioral phenotype of autism may result from research on mediators of social behavior, which may also assist discover the neural networks necessary for typical social interaction. The core approach techniques for modelling ASD in animals are explained and examined in this article, along with the behavioral strategies used in the research among these models.
Autism is an major neurological problem marked by repetitive or stereotyped behaviours, anxiety, and cognitive dysfunction, as well as neurodevelopmental abnormalities in social communication1,2. Although the cause of Autism is poorly understood, research suggests that both a significant hereditary component and environmental factors work together as triggers3,4. The diverse collection of diseases known as autism is typified by issues with verbal and Nonverbal expression, social interaction, and recurring behaviours brain's circuitry connections and It has been discovered that architectural organization has a significant role in the start of symptoms, and the Autism cause is thought to begin earlier in development5.The following section will go over behavioral symptoms of ASDs, such as repetitive and stereotypical behaviours, anger and irritability, Excessive activity and lack of focus, and Social disability. Stereotyped motor actions like tapping, wiggling, twirling or flailing the hands are examples of repetitive behaviours. They might also include rigid adherence to rituals or routines that don't work. Because it can be challenging to differentiate these symptoms from those of obsessive-compulsive disorder (OCD), this review will cover both conditions' treatments. Severe outbreaks of rage and impulsive hostility toward oneself or others are examples of irritability in ASDs. It is estimated that up to Thirty percent of kids and teenagers with ASDs feel lightly to serious irritation6. Between 15 and 25 percent of autism cases are connected to recognizable hereditary conditions (such as neuralgia and a tuberous, Rett's illnesses, and the fragile X chromosome) as well as viral illnesses such congenital rubella or metalogical pathologies7. Although hyperactivity and inattention are common in individuals with ASDs, an assessment of ASD prevents a related assessment of attention deficit hyperactivity disorder, according to the forthcoming Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition, Text Revision (DSM-IV-TR) criteria. According to estimates, between 40% and 59% of kids with ASD diagnoses also meet the requirements for ADHD8,9. The 3rd edition of the treatment and Statistical Manual of Mental Disorders-III published by the APA in 1980 added autism as a subgroup under the diagnostic category of pervasive developmental illness. This was done to communicate the idea that a more extensive range of conditions are included in behavioral deficiencies. Four categories of PPD were identified development in childhood PDD, legacy autism, fragile autism, and an uncommon type Ten. In the 1980s, Wing and Gould placed autistic children on an escalator with other abnormal children by defining autism in terms of habits rather than psychosis11,12. The notion that Kids with autism lacked " theory of mind," or the the capacity to identify one's own psychological conditions and other individuals, It is a vital part of the Socializing experience and was developed in the middle to late 1980s as a result of study by Alan Leslie, Uta Frith, and Simon Baron-Cohen13. Autism was first recognized as a disability in 199014. However, it is still unclear how exactly calretinin (CR) regulates neuronal excitability and the behavioural abnormalities associated with ASD. Several lines of evidence link the use of antiepileptic medications, such as VPA, throughout being pregnant to increased Autism hazards in the fetus15. Access to VPA during pregnancy caused Attributes unique to men in synapse formation, morphological growth and societal participation, whereas female progeny mainly displayed minor abnormalities16. It was suggested that vaccinations for babies could contribute to the development of autism; however, this claim is still unproven17. for researchers to Search into the underlying causes of Autism occurrences and to Build medications and Alternative approaches for treating the symptoms of these conditions Autism model animals are being designed.
Behavioural Assessments for Analysing ASD Models
As autism models for animals, several behavioural test methods are employed to estimate the outward signs of abnormalities. The two universal classical tests (Morrison, open field, and three chamber, radial maze, water maze, T-like maze), and techniques for identifying particular A three-section apparatus for measuring social communication, analysis, and reproducing ultrasonic indicators in social interaction investigations, and other characteristics of autism disorders are detailed below.
The behaviour of the animals is often monitored in settings where they are regularly (familiarly) housed, right in the cages where they typically reside. It is convenient to combine and modify specific procedures, which enables experimenters to provide a more thorough and in-depth assessment of the phenotypic manifestations in the corresponding models. Anxiety, cognitive flexibility, and social behaviour are typically examined in a battery of techniques used to evaluate Autism models for animals. The usefulness of estimating anxiety levels in this situation is questionable, though, as variations in this index are not associated with certain traits of autism. For this reason, it is more practical to focus on the first two lines of inquiry: the analysis of cognitive process flexibility and shifts in animal social interactions18,19.
Table 1. Animal Behavior Tests to evaluate Autism
|
Sr. No |
Behavioural Domain |
Test |
Observation |
References
|
|
1. |
Social Interaction |
Three-chamber social approach test - Social sniffing test |
Reduced time spent interacting with other animals |
19,20,21 |
|
2. |
Communication |
Ultrasonic vocalization (USV) test in pups |
Lack or abnormal calls |
19 |
|
3. |
Repetitive Behaviour |
Marble burying test, self-grooming |
An increase in repeated activities |
19 |
|
4. |
Cognition, Memory |
Morris water maze, Y-maze |
Memory and learning impairment |
19 |
|
5. |
Anxiety |
Open field test, elevated plus maze |
Exceptional reactions to anxiety |
22,19 |
Test of three-chamber social interaction
The apparatus has three chambers of 40 × 22 × 20 cm, each with openings measuring 5 × 8 cm to facilitate exploration. The examination was conducted within limited illumination (5 lx). The illumination in each a compartment was examined using a handheld lux meter. Unlike the study mice, the animals selected as the foreign outsiders had been selected for breed, lifespan, and gender. The animal in question was placed in the middle area that is vacant during the habituation phase and given ten minutes to explore the entire room. During the sociability phase, A metal box that is vacant (new item) was put in the opposite room, while a wire cage (10 cm in diameter) containing a chosen new animal (visitor 1) was put in the opposite room. To get rid of side preference, the stranger and unfamiliar object were gradually moved to either in the opposite direction. The test animal was given the freedom to investigate the new environment for ten minute.
An unidentified prominent animal (visitor 2) was placed in the canter zone for the uniqueness of society preference test on the right side of the wire cage. For ten minutes, the subject mouse was once more free to roam around each compartment. Time stayed in each container was noted, as was the duration of time stayed detecting or connecting with an unidentified animal. Every time an animal's nose came within two centimetres of an unknown animal or touch its side a compartment or tail, the probing period was observed.
To assess friendliness, we determined each mouse's discriminating index (the period of time expended with a new mouse minus time expended with an unfamiliar object)) / (total amount of period expended using the stranger object and novelty the mouse 1). The following formula was used to determine the discrimination index: To determine preference for social novelty, divide The duration of time expended with a new mouse (1) divided by the period of time expended with an unfamiliar mouse (2).20,21
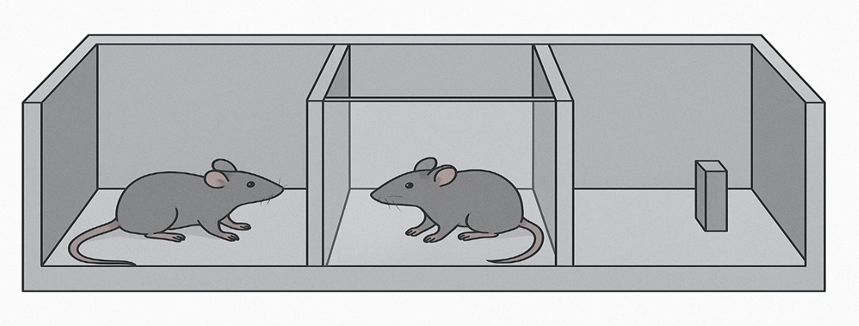
Figure 1. Test of three section social communication
Open-field (OF) test
A mice's locomotor movements and anxiety-like behaviours were described using the OF test. Theoretically, a 40 x 40 cm A central field was separated from the white plastic OF compartment (20 × 20 cm) and a periphery a space for research. At the beginning of each test, each mouse was positioned in the canter of the OF and given six minutes to discover the space. The length and a total amount of participants travelled in the OF were recorded, together with the duration and length travelled in the central and expanded arenas. Limelight Software was applied to automatically observe the mouse locations.22
The Elevated plus maze (EPM) test
EPM- test apparatus, which has two open arms (30 × 5 cm) and two closed arms (30 × 15 × 5 cm) that intersect at a 90° angle to form a plus, is attached by a main pillar. A mouse was positioned in the middle of the device, in the open arm looking, and given ten minutes to be wander around. Both The overall quantity of open arms records and the duration of open arms it was determined. Between each test, the equipment was dry after cleaning with 75 percent ethanol.
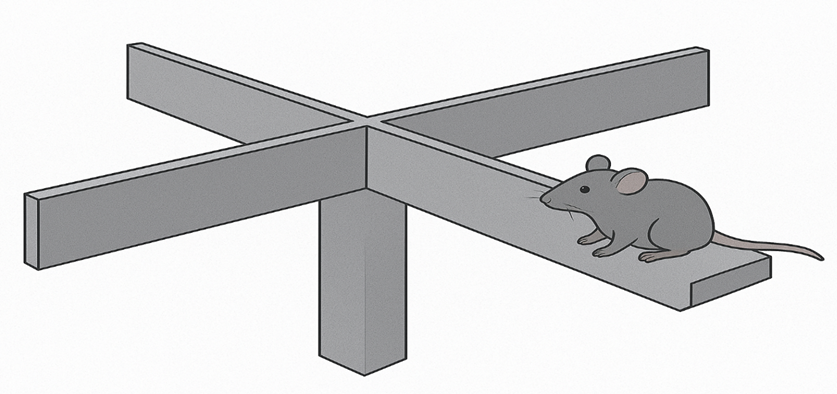
Figure 2. EPM- test apparatus
Morris water maze (MWM) test
A MWM test (diameter = 120 cm) with white water (23 ? C) and many Graphic hints were utilized to assess memory and spatial learning skills. The mice had to create distal additional maze indicators to locate undercover frame over the five-day training sessions. During a period of five days, they participated in four tests (60-s cut-off) / day with a five- minute break delay after being brought in from one of four possibilities in a daily selected sequence. In order to locate the platform, the swimming velocity, route, and latency were captured with a video observation device from Xinruan. On day six, a 60-second a probing examination was used to assess spatial cognition after the platform was taken out of the water. The goal segment was where the elevated area is situated. Probe test performance was measured by the quantity of entries and the proportion period expended in the intended compartment. In order to assess visuomotor impairments, the visible platform test was conducted on day 7. The rodents possessed one chance to trace an elevated platform that was visible and signalled by a cue, with a 60-second cut-off. The latency of the platform's navigation speed was measured using a video-tracking system.
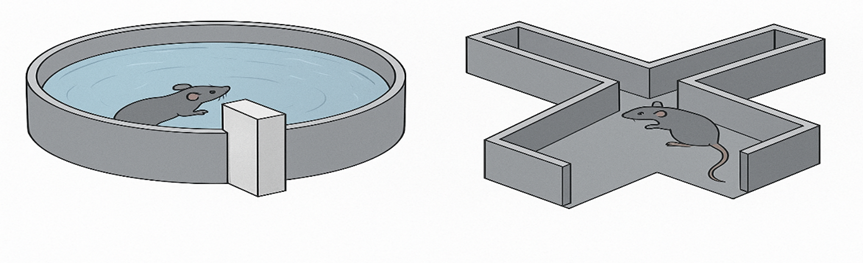
Figure 3. MWM- test Figure 4. Y- maze test
Transmission electron microscopy (TEM)
The nerve cells was ready for microscopic examination23,24. After inducing severe unconsciousness, Mice received an ice-cold perfusion. 0.9 percent biological salinity, 4 percent paraformaldehyde and 2 percent glutaraldehyde in 0.1 M PBS (pH 7.4). After that, the hippocampal tissue was isolated and applied with paraformaldehyde at 4 percent for an entire night at 4°C. The specimens were processed fixation, 1 percent of osmium tetroxide after fixing for 1.5 hours, dehydration in ethanol (There was 1 percent uranyl acetate in the 70 percent ethanol), and implantation in epoxy resin. For the extremely thin segments, The portions that are semithin were given at 500 nm Depths, 1 percent toluidine blue stain, and seen in the light microscope. An ultra-cut Leica ultra small microscope (EM UC7, Leica, Germany) was used to cut ultrathin sections (90 nm) of the appropriate area. Following staining with lead citrate and uranyl acetate, these sections were inspected applying an electron microscopy (Tecnai G2 20 TWIN, FEI, USA). Postsynaptic density and synaptic vesicles were used to identify synapses. We examined 60–80 electron microscopic pictures on average for each group. As previously mentioned25 we quantified the following parameters: synaptic active zone length, strength of synaptic thickness, breadth of the synapse cleft, and synapse density per 100 μm 2.
Analysis of statistics
For statistical evaluation, The GraphPad Prism 26.0 (GraphPad Software) was utilized. The mean ± SEM is used to display the information. The Tukey's multi compare technique post hoc comes after the Unpaired One-way ANOVA or Student's t-test, were employed to assess the data. for the behavioural assessment outcomes. Repeated actions The findings because the electrical reactions to higher current administrations were significantly examined using ANOVA. When p < 0.05, a difference was deemed Significant statistically.
Investigations into Cognitive Rigidity and Interest Limitations
Cognitive rigidity is autism's second key symptom18. suggested utilizing the T-maze to assess this factor. After teaching the animal to locate a meals incentive in a subdivision, the prize was moved to the opposing arm. The speed at which the animal alters its behaviour and starts searching for food in a new location is noted by the experimenters. Other possible approaches include calculating how long it takes to find the intended level in the MW-maze after it is removed and estimating the rate at which the conditioned reflex attenuates when reinforcement is removed.
an overabundance of common behavioral behaviour, such grooming, maintaining oneself upright, shaking, whirling, flying, etc., are examples of rodents' repetitive and stereotypical pattern of behaviour26,27,28. Usually, during open field tests or during regular behaviour, these behavioral aspects are seen and documented right in the animal cages where the animals are housed. Usually, during open field tests or during regular behavior, these behavioral aspects are seen and documented right in the animal cages where the animals are housed.
An intriguing method for examining the stereotypical pattern behaviours in models of ASD-specifically, in BTBR--altered mice-is the "burying-stones" test, also known as the "marble burying test"29.
Animals are placed with a thick coating in the exam field of matting for this test. It is Animals initially acclimated to the place in order to reduce the tension response to the unfamiliar surroundings. Tiny gravel, pieces of glass, or plastic pellets are placed on the bed by the experimenters, who then count how many of these objects the test animal excavates. One common example of Guarding actions in mice, specifically mice and rats, is the reaction of burying unknown objects30.The animal tries to hide anything it believes to be disagreeable and hazardous from the ground or material that expands with its muzzle and forelimbs. In the natural world, these could be snakes, insects, or scorpions; in the lab, they could be stimulating shock electrodes. Bits of food, they stone, and glass beads—items that aren't meant to be harmful—are also easily buried. One of the main characteristics of ASD in humans is recurrence of behavior, which can be well-indicated by the repeated, obsessive nature of such a behavioral pattern in models of the disorder. Because anxiolytics (like diazepam) significantly reduced the severity of these responses, burying the stones was once thought to be an anxiety correlate31. However, such conduct is currently thought to reflect obsessive/compulsive characteristics and be primarily tied to the novelty of objects. For this reason, this test is currently accepted in models of obsessive-compulsive disorders and autism 32.
The test settings, which involve placing the animal in a chamber with numerous burrow-like entrances in the walls and bottom, can cause the hypothesized restriction of interest in rodents to appear. Rats in good health closely inspect various holes, sniffing them and inserting their muzzles into these "burrows." The same "hole" (or several "burrows") is greatly preferred by the animals exhibiting autism symptoms, who repeatedly explore them19.
Animals are frequently asked to choose between two branches in a Y-shaped maze when their behavior is being examined, and neither branch receives food incentive.
After visiting, say, the left arm, healthy rodents typically go on to the right. One could assume that a propensity to repeatedly visit the same arm would be seen in autism models. However, this test does not identify cognitive rigidity and most likely describes the specialization of research activity33.
Modelling and investigation of animal communication deficiencies
Since only humans use voice for communication, it might be challenging to replicate the third element of the "autistic triad" in animals. As a result, we can only identify the associated communication problems in humans. A However, it is known that animals can use a variety of signalling systems to build communication relationships. It was discovered that rodents (hamsters, rats, gerbils, mice etc.) consistently communicate using ultrasonic auditory signals34. These indicates were seen in almost every aspect of the rats' social lives, including socialization, violent sexual activity, and more. It has been demonstrated that children who attempt to find the nest produce ultrasonic waves that enable their mothers to locate them and bring them back to a safe area35. The female left the nest and started looking after hearing the signal. Nevertheless, this mother did not respond to lifeless or drugged newborns that did not show these signs. The baby mice's ultrasonic pulses were utilized36. When the equivalent observe was activated in an vacant chamber of the study setup using the nest in place, most of the moms relocated to the previously described chamber, that was totally absent of newborn mice, plush animals of a suitable size, and the matching fragrance tracks. Rats were used in similar investigations later on. Therefore, compared to visual and olfactory inputs, ultrasonic vocalization (USV) likely has a far greater influence on mother behavior in rodents.
Models of autism diseases show significant disruptions of USV when compared to the vocalization of healthy animals. Specifically, when separated from their moms and other young animals, Shank1–/–mutant mice's progeny generated lower signals37. Vocalization problems were also evident in mature model animals. When given the opportunity to sniff female urine, mature male strains of mice BTBRT T+tf/J (BTBR) generated fewer ultrasonic indications and departed from fragrant tags than animals of the unconventional style38.
Vocalization Problems in songbirds were also studied39. It is thought that these birds learn unique songs that are exclusive to particular species throughout their lives in addition to using vocalization for communication. That can be mentioned that just men are capable of "sing" corresponding tunes, and their rivalry in a female partner of breeding heavily relies on his performance. That's believed it human speech learning impairments are caused by mutations in the same gene, FOXP240. rodents' vocalization41,42 and songbirds' songs43. It was demonstrated that learning songs was hampered by "switching off" this gene due to lentivirus effect44. It is well known that songbirds pick up a species-specific tune by imitating an older bird, or "teacher." Birds that had problems with the FOXP2 gene either reproduced the song incompletely or with considerable distortion.
When competing for a female, the male bird did not benefit from such aberrant vocalization. It cannot be ruled out that studying the characteristics of bird vocalization can provide some knowledge about modelling illnesses similar to autism.
Strategies for Repetitive Activity and Special Interests
Repetitive activity in mice can involve persistent sniffing, circling, burrowing, or constant running and jumping in the cage45,46,47. Repetitive grooming and scratching can result in skin lesions and ear injuries48. Both open-field testing and home-cage monitoring can reveal aberrant stereotypies.
Excessive devotion to routine is another manifestation of autism's propensity for repeated behavior. Reverse studying in the MW-Maze challenge may be applied to Calculate resistivity to changing a knowledgeable behavioral arrangement. Mice are educated to find a hidden climb framework that is partially submerged in a circular pool of milky liquid in this exercise. Mice develop swimming abilities straight to the hiding ledge from any access spot on the pool's edge during a few days. Prior to an end test, the platform for escaping is removed to evaluate the task's spatial learning. Animals typically swim in circles. over the platform's placement. The escape platform is moved to a distinct area of the pool in the subsequent testing phase, and the mice's the newly established location's learning capacity is reassessed. Inadequate or delayed formation in the context of reversal studying may act as an example of mental stiffness similar to autism. According to this theory, Fmr1-null mice demonstrated adequate visual education acquisition in the MW-maze test but inadequate Acquiring knowledge while reversing stage49,50. The water maze test may not be possible for Mice with impaired vision or motor skills. In these situations, learning acquisition and retrieval can be assessed using a straightforward T-maze experiment. In order to receive a food reward, Mice are taught to go in either the left or right arm of the labyrinth. When a significant number of right decisions are noted, the reinforcement arm is switched, requiring the mouse to choose the other arm in order to receive rewards. One drawback of this duty is that I have to go without eating before acquisition, which may result in problems with motivation levels among strains. The spontaneous alternation test is another method that makes use of the T-maze. It is predicated on mice's innate propensity to switch between arms in a maze51. Exploration metrics on a hole board, such as the number of nose pokes into a certain collection of holes relative to the entire board52,53, and the inspection of unfamiliar things in an open field are other methods to evaluate repeated behavior54,55.
CONCLUSION
ASD is a complex neurological condition that impacts communication, social interaction, and repetitive behaviours. Environmental and genetic variables both have an impact. Its pathophysiology is impacted by genetic abnormalities like fragile X and Rett's as well as prenatal exposure to substances like valproic acid. Developing successful therapy approaches and interventions requires an understanding of ASD through behavioral and animal models. It is not possible to identify ADHD alongside an ASD diagnosis. animal models of autism, specific behavioral testing procedures (Morrison, open field, and three chamber, radial maze, MW-maze test, T-maze) and ways for identifying specific A Social contact within 3 compartments tool measurement. The T-maze, MW-maze test, and marble burying exams are examples of a tasks that can be used to simulate and assess cognitive rigidity and repetitive behaviours, two fundamental characteristics of autism. These paradigms evaluate the animal's capacity for exploration, adaptation, and recurrent or compulsive activity. These assessments provide important insights into behavioral phenotypes similar to ASD by differentiating anxiety-related from obsessive behaviours. All things considered, these models are crucial resources for researching the neurological processes and treatment approaches in autism. Modeling the communication deficits of autism in animals is challenging, yet studies of ultrasonic vocalizations (USVs) in rodents and song learning in birds offer valuable insights. Rodents use USVs for social communication. Similarly, mutations in the FOXP2 gene in songbirds cause abnormal song learning, paralleling human speech difficulties. These findings suggest that vocalization studies can illuminate the neural and genetic bases of autistic communication deficits. Repetitive behaviors and cognitive rigidity, another hallmark of ASD, can be modeled through behavioral tests like the Morris water maze, T-maze, and hole-board assays. These paradigms reveal persistence, reduced flexibility, and stereotypic patterns in ASD models. Together, such animal models provide crucial platforms for understanding autism’s core features and testing potential therapeutic interventions.
This paper is a review and has nothing to do with any experimental research.
REFERENCES



Ranjeet Kumar, Mudit Kumar, Dr. Yogesh Chand Yadav, Autism Spectrum Disorders Related Mouse Behavioral Approaches, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 1, 707-718. https://doi.org/10.5281/zenodo.18186331
 10.5281/zenodo.18186331
10.5281/zenodo.18186331
