Sigma Institute of Pharmacy, Ajwa Nimeta Road Bakrol, Vadodara- 390019, Gujarat, India
Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and its associated proteins have revolutionized modern biology. Originally discovered as an adaptive immune system in prokaryotes, CRISPR–Cas technologies have evolved into highly versatile genome-editing tools. The development of base editing and prime editing has extended the precision and scope of genetic modifications, avoiding double-strand breaks and allowing correction of pathogenic mutations at single-nucleotide resolution. Complementary computational platforms such as STITCHR provide systems-level insights into protein–chemical interactions, supporting rational therapeutic development and drug repurposing. This review summarizes the historical development of CRISPR, its molecular mechanisms, generational advancements, and innovations in delivery systems. We further examine clinical applications across oncology, liver, cardiovascular, and hematological disorders, highlight CRISPR-based diagnostics, and discuss integration with bioinformatics platforms including STITCHR. Ethical, social, and regulatory considerations are critically analyzed in the context of somatic and germline editing. Finally, we discuss future prospects where CRISPR and computational systems biology converge to advance precision medicine. This review comprehensively evaluates CRISPR and STITCHR, providing perspectives on their transformative potential in genetic engineering and therapeutics.
The capacity to manipulate genomes with unprecedented precision has dramatically reshaped the biological sciences, biotechnology, and medicine. Genome-editing technologies represent one of the most significant scientific breakthroughs of the 21st century. Initially dominated by zinc-finger nucleases (ZFNs) and transcription activator-like effector nucleases (TALENs), the field rapidly shifted to CRISPR–Cas systems due to their simplicity, adaptability, and robustness. (1,2) Unlike ZFNs and TALENs, which require laborious protein engineering, CRISPR relies on a short guide RNA that directs a nuclease to a target site, offering programmability at low cost. (3)
The origins of CRISPR lie in the adaptive immune system of bacteria and archaea, where it provides resistance against invading phages and plasmids. Since the pivotal 2012 demonstration of CRISPR–Cas9 as a programmable editing tool, (4) research has exploded across multiple domains. The award of the 2020 Nobel Prize in Chemistry to Emmanuelle Charpentier and Jennifer Doudna recognized this milestone. (5)
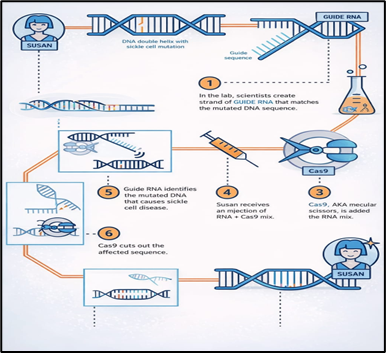
Figure 1 Introduction to CRISPR
The development of derivative technologies such as base editing (6) and prime editing (7) extended CRISPR’s capabilities beyond double-strand break (DSB) repair, allowing nucleotide-level precision. These tools enable correction of single base mutations without introducing DSBs, potentially addressing ~89% of known pathogenic variants (7).
Simultaneously, computational approaches have expanded to contextualize genome editing. STITCHR (8), a bioinformatics platform mapping protein–chemical interactions, offers systems-level understanding, aiding therapeutic discovery and drug repurposing. This synergy between experimental genome editing and computational biology heralds a new era of precision medicine and integrative biotechnology.
2. HISTORICAL DEVELOPMENT OF CRISPR
The origins of CRISPR trace back to fundamental observations in microbiology. In 1987, Ishino and colleagues (9) reported the presence of unusual repetitive DNA sequences in Escherichia coli, although their biological role was not understood at the time. This mystery persisted until Mojica and co-workers (10) proposed that these repeats were derived from invading genetic elements, functioning as a form of molecular memory that enables microbial defense. Definitive experimental evidence came in 2007, when Barrangou et al. (11) demonstrated that CRISPR loci confer adaptive immunity in Streptococcus thermophilus by protecting against bacteriophage infection.
A pivotal breakthrough occurred when Charpentier and Doudna reconstituted the CRISPR–Cas9 system in vitro, showing that the Cas9 nuclease could be guided to specific DNA sequences by a programmable dual-RNA complex. (4) This finding revealed an unexpectedly simple and versatile mechanism for sequence-specific DNA cleavage, fundamentally reshaping genome engineering. Soon after, independent studies by Zhang and colleagues (12) and by Church’s group (5) confirmed that CRISPR–Cas9 could function efficiently in mammalian cells, marking the beginning of the modern CRISPR genome-editing era.
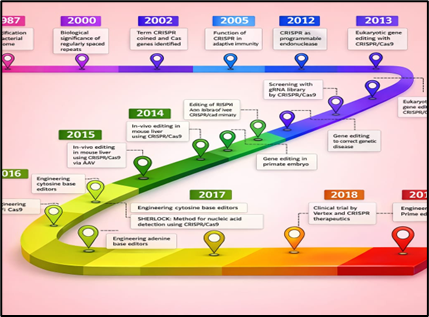
Figure 2 Historical Development of CRISPR
In the years that followed, the diversity of CRISPR systems expanded rapidly. Cas12a (formerly Cpf1) was identified as an alternative DNA-targeting nuclease with distinct PAM requirements and staggered DNA cleavage patterns. (13) Cas13 was subsequently characterized as an RNA-targeting effector, opening new avenues for transcriptome manipulation and CRISPR-based diagnostics. (14) More recently, the discovery of compact nucleases such as Cas14 and CasΦ, derived from bacteriophages, further reduced protein size constraints and offered potential solutions to delivery challenges. (15) Collectively, these advances have greatly broadened the CRISPR toolbox, enabling precise manipulation of DNA, RNA, and epigenetic states.
3. MECHANISM OF CRISPR–CAS9
CRISPR–Cas9 functions as an RNA-guided endonuclease system in which a single-guide RNA (sgRNA), formed by the fusion of CRISPR RNA (crRNA) and trans-activating CRISPR RNA (tracrRNA), associates with the Cas9 protein to direct it toward a complementary DNA sequence located adjacent to a protospacer adjacent motif (PAM). (16)
In the case of Streptococcus pyogenes Cas9, this PAM sequence is most commonly NGG. Following accurate target recognition, Cas9 introduces a site-specific double-strand break (DSB) in the DNA. The cellular response to this break is governed by endogenous repair mechanisms, primarily non-homologous end joining, an error-prone pathway that frequently generates insertions or deletions and is widely exploited for gene disruption, and homology-directed repair, a template-dependent pathway that enables precise sequence modification when an exogenous donor template is provided. (17)
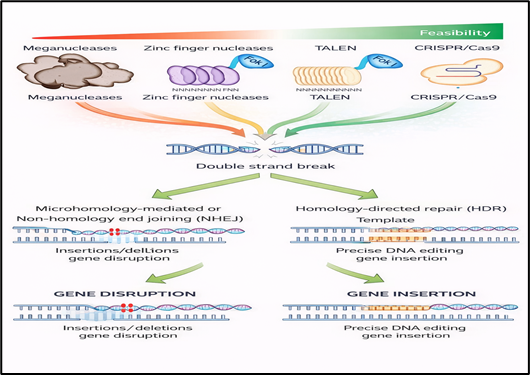
Figure 3 Mechanism Of CRISPR
To enhance editing precision and minimize off-target effects, several engineered Cas9 variants have been developed, including nickase Cas9, which cleaves only a single DNA strand to reduce genotoxicity, high-fidelity Cas9 variants such as eSpCas9 and SpCas9-HF1 that exhibit improved mismatch discrimination, (18) and PAM-relaxed variants such as xCas9 and SpCas9-NG that expand the range of editable genomic sites. (19) This refined mechanistic understanding and ongoing protein engineering efforts laid the foundation for the development of base editing and prime editing technologies, which overcome key limitations associated with double-strand break–dependent genome editing.
4. GENERATIONS OF GENOME EDITING
Genome-editing technologies have progressed through four major developmental phases, reflecting increasing precision and ease of use. The first generation comprised zinc-finger nucleases, which combined engineered zinc-finger DNA-binding domains with the FokI endonuclease to achieve targeted genome modification; however, their application was limited by the complexity and labor-intensive nature of protein design. (20)
This was followed by transcription activator-like effector nucleases, derived from plant pathogen TALE proteins, which offered improved modularity and targeting flexibility but still relied heavily on extensive protein engineering. (21) The third generation emerged with CRISPR–Cas9, a transformative RNA-guided system that exploits RNA–DNA complementarity for programmable genome editing, enabling scalability and widespread adoption across biological research and therapeutic development. (4,5)
The most recent, fourth-generation tools include base editors and prime editors, which enable highly precise genetic modifications without introducing double-strand breaks and allow direct correction of pathogenic variants at single-nucleotide resolution. (6,7) Collectively, this evolution illustrates a fundamental shift from protein-centered genome-editing platforms to RNA-guided technologies, culminating in tools capable of unprecedented precision.
5. BASE AND PRIME EDITING
Base editors have emerged as a major advance in precision genome engineering by enabling targeted nucleotide conversions without introducing double-strand breaks. Cytosine base editors (CBEs) achieve C→T transitions through the fusion of a nickase Cas9 (nCas9) with cytidine deaminases, (6) while adenine base editors (ABEs) mediate A→G conversions by harnessing engineered tRNA adenosine deaminases. (22) Although these systems eliminate the need for donor DNA templates and reduce the risks associated with double-strand breaks, they are limited by issues such as bystander editing within the editing window and unintended off-target deamination events (23).
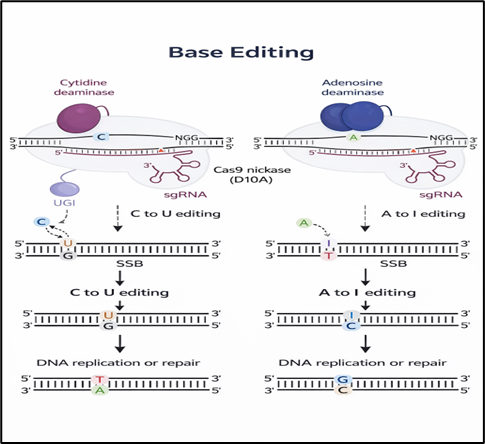
Figure 4 Base Editing
Prime editing, introduced in 2019, further expanded the genome editing toolkit by integrating nCas9 with a reverse transcriptase and a prime editing guide RNA (pegRNA) that simultaneously specifies the target site and encodes the desired genetic modification. (7) This versatile platform enables all twelve possible base substitutions, as well as precise small insertions and deletions, allowing correction of a broad range of pathogenic variants. While prime editing offers greater flexibility compared with base editing, its clinical translation is still challenged by suboptimal efficiency and delivery constraints. Ongoing improvements in pegRNA design, along with the incorporation of machine learning–based optimization strategies, are progressively enhancing editing performance and reliability. (24)
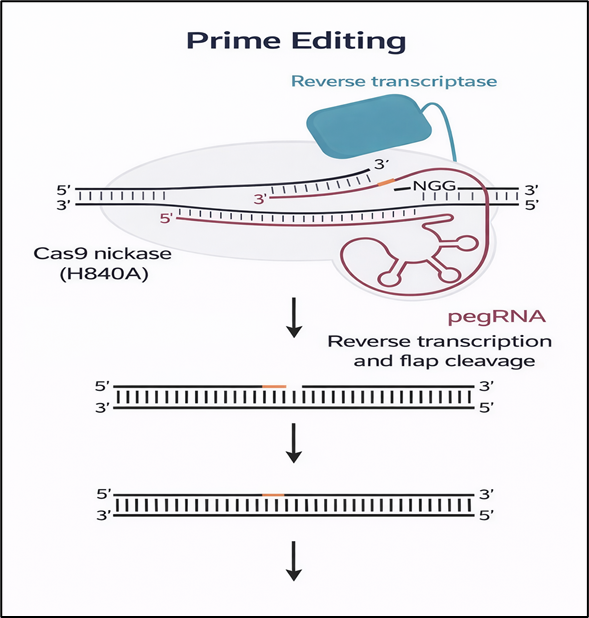
Figure 5 Prime Editing
6. DELIVERY SYSTEMS
Delivery remains a major bottleneck in the clinical translation of genome editing technologies. Viral vectors continue to be extensively explored, with adeno-associated viruses (AAVs) favored for in vivo applications, although their clinical utility is constrained by a limited cargo capacity of approximately 4.7 kb and concerns related to immunogenicity. (25) Lentiviral vectors enable stable genomic integration and long-term expression but carry inherent risks of insertional mutagenesis and oncogenesis. In parallel, nonviral delivery platforms have gained momentum; lipid nanoparticles (LNPs), successfully validated through mRNA vaccines, have demonstrated strong potential for CRISPR-based therapeutics, while polymeric carriers and gold nanoparticles are also emerging as promising alternatives. (26,27)
Exosomes represent another attractive strategy due to their biocompatibility and intrinsic targeting capabilities. (28) Additionally, physical delivery methods such as electroporation, microinjection, and hydrodynamic injection are highly effective in ex vivo settings, particularly for cell-based therapies. (29) To overcome the limitations of individual systems, current research is increasingly focused on hybrid delivery platforms that integrate the high efficiency of viral vectors with the improved safety profiles of nonviral approaches, alongside the development of compact Cas variants to further ease delivery constraints.
7. CLINICAL APPLICATIONS
CRISPR technologies have progressed rapidly from experimental tools to clinical applications across a wide range of diseases. In oncology, genome editing has been employed to improve CAR-T and TCR-T cell therapies by targeting oncogenes such as TP53 and NOTCH1 as well as immune checkpoint regulators including PD-L1. (30–32) In the context of liver disorders, base editing has successfully restored enzyme activity in preclinical models of phenylketonuria, (33) and clinical studies in transthyretin amyloidosis have demonstrated the feasibility and efficacy of in vivo CRISPR-mediated gene editing. (34) Cardiovascular research has also benefited from these advances, with CRISPR-based editing of PCSK9 leading to significant reductions in cholesterol levels in preclinical models, highlighting its potential for the treatment of atherosclerosis. (35) In hematological diseases, particularly sickle cell disease and β-thalassemia caused by point mutations in the HBB gene, CRISPR-edited hematopoietic stem cells have shown promising therapeutic outcomes, with prime editing offering the possibility of direct and precise mutation correction. (36–38) Beyond these areas, CRISPR-based interventions are being explored for ophthalmic conditions such as Leber congenital amaurosis, neuromuscular disorders including Duchenne muscular dystrophy, and infectious diseases such as HIV.
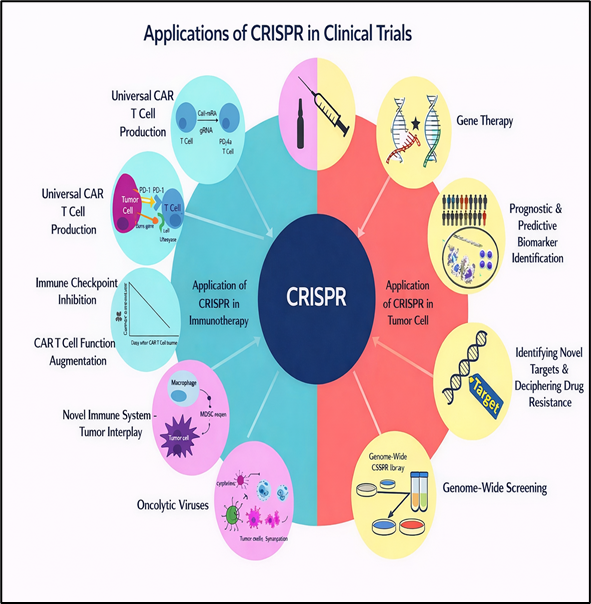
Figure 6 Clinical Applications of CRISPR
8. CRISPR IN DIAGNOSTICS
CRISPR technology has been adapted for diagnostic applications by harnessing the collateral cleavage activity of Cas12 and Cas13 nucleases. The SHERLOCK platform, which is based on Cas13, allows highly sensitive detection of RNA targets, (39) whereas the Cas12-based DETECTR system is designed for the identification of DNA pathogens. (40) These diagnostic approaches offer rapid, portable, and cost-effective testing solutions and played a significant role in large-scale surveillance efforts during the COVID-19 pandemic. As such, CRISPR-based diagnostics hold considerable promise for expanding access to reliable testing, particularly in low-resource and decentralized healthcare settings.
9. BIOINFORMATICS INTEGRATION: CRISPR AND STITCHR
STITCHR integrates protein–chemical interaction networks to provide a systems-level context for CRISPR-based interventions, enabling a deeper understanding of how edited genes interface with pharmacological pathways. (8) By connecting genome-edited targets with curated drug and chemical interaction databases, STITCHR supports drug repurposing strategies, facilitates off-target assessment, and informs rational therapeutic design at the network level. Additional complementary bioinformatics resources further strengthen this integrative framework, including STRING for mapping protein–protein interaction networks, (41) DrugBank for comprehensive drug–target annotations, (42) and machine-learning–based CRISPR off-target prediction tools that aid in the design of highly specific guide RNAs. (43) Collectively, the integration of CRISPR datasets with computational systems biology platforms advances precision medicine by enabling informed, data-driven therapeutic development.
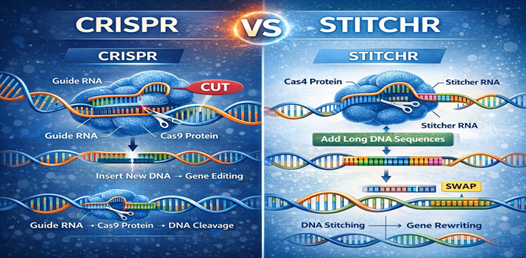
Figure 7 CRISPR VS STITCHR
10. ETHICAL, SOCIAL, AND REGULATORY ISSUES
The ethical discourse surrounding CRISPR technologies intensified markedly after He Jiankui’s 2018 announcement of genome-edited embryos intended to confer resistance to HIV, an event that raised profound global concerns regarding the permissibility of germline genome editing. (44) This episode brought critical ethical challenges to the forefront, including safety issues such as mosaicism, unintended off-target effects, and unknown long-term consequences; questions of equity related to fair and global access to CRISPR-based therapies; dual-use concerns involving potential misuse for human enhancement or biotechnological harm; and substantial regulatory disparities across countries, prompting calls for cohesive international oversight and governance frameworks. (45,46) While somatic genome editing for therapeutic purposes has gradually gained cautious scientific and regulatory acceptance, germline editing continues to be broadly restricted or prohibited in most jurisdictions worldwide.
11. FUTURE DIRECTIONS
Future directions in genome-editing research are focused on overcoming current technical and translational limitations through coordinated molecular and computational innovations. These include the engineering of compact Cas proteins to facilitate efficient and safe delivery, particularly for in vivo therapeutic applications. (47) Parallel efforts are directed toward the development of next-generation base editors with minimized bystander and off-target editing to enhance precision and genomic safety. (48) In addition, prime editing efficiency is being improved through artificial intelligence–driven design of prime editing guide RNAs (pegRNAs), enabling higher editing accuracy and broader applicability across diverse genomic contexts. (49) Importantly, the integration of genome-editing technologies with advanced bioinformatics platforms such as STITCHR, coupled with artificial intelligence, is enabling systems-level therapeutic design by linking edited genetic targets with drug interaction networks, thereby accelerating precision medicine and rational therapeutic development. (50) The convergence of genome editing, nanomedicine, AI, and bioinformatics will define the next decade of therapeutic innovation.
12. CONCLUSION
CRISPR–Cas9, base editing, and prime editing constitute transformative tools in modern biology. Their integration with bioinformatics platforms such as STITCHR elevates genome editing from site-specific interventions to systems-level therapies. As delivery, efficiency, and ethical challenges are progressively addressed, CRISPR technologies will play a central role in next-generation medicine and biotechnology.
REFERENCES

Vidhi Patel, CRISPR vs STITCHR: Comprehensive Review on Base Editing, Prime Editing, and Therapeutic Applications, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 1, 785-795. https://doi.org/10.5281/zenodo.18198339
 10.5281/zenodo.18198339
10.5281/zenodo.18198339
