Womens College of Pharmacy, Peth Vadgaoan.
Valeriana jatamansi Jones (synonymously known as Indian valerian or Tagar) is a perennial herb indigenous to the Himalayan region and used traditionally across Ayurvedic, Unani and Chinese systems for central nervous system (CNS) disorders and allied indications. This review surveys the phytochemical landscape of the species including valepotriates (e.g., valtrate, didrovaltrate, acevaltrate), sesquiterpenes (e.g., patchoulol, jatamansone, ?-caryophyllene), flavonoids and phenolics (e.g., quercetin, apigenin, gallic acid), alkaloids (e.g., actinidine, valerine), essential oils (e.g., valeranone, bornyl acetate) and additional lignans/fatty-acid glycosides and collates their pharmacological activities. Extracts and isolated compounds of V. jatamansi display a range of biological effects: CNS modulation (sedative, anxiolytic, anticonvulsant, neuroprotective), antioxidant and anti-inflammatory actions, antimicrobial/antifungal effects, anticancer cytotoxicity, cardioprotective/hepatoprotective influence, endocrine/metabolic regulation, and other adaptogenic/spasmolytic/gastroprotective activities. Mechanistically, data indicate involvement of GABAergic modulation, monoamine oxidase (MAO) inhibition, ion-channel effects (Calcium V channels), oxidative-stress defense (SOD, catalase, glutathione peroxidase), neurotrophic factor up-regulation (e.g., BDNF, NGF) and PI3K/Akt signaling especially in neurodegeneration contexts. Toxicity and standardization issues (unstable valepotriates, essential-oil variability) remain a challenge; acute and sub-chronic animal studies indicate favorable safety margins though human clinical data are scant. Recent advances (2020–2025) include nanocarrier formulations, network-pharmacology/omics approaches and improved cultivation/conservation strategies. For translation into neurotherapeutic herbal products, future research must emphasise rigorous clinical trials, pharmacokinetics, biomarker-driven mechanism studies, and standardised extract development.
The genus Valeriana (family formerly Valerianaceae; more recently placed in Caprifoliaceae/Valerianoideae) comprises approximately 200 species of perennial herbaceous plants distributed across temperate regions of Eurasia and North America (Ma et al., 2021; Li et al., 2022). Among these, Valeriana jatamansi Jones (synonym Valeriana wallichii DC.) commonly termed Indian valerian or Tagar occupies an important place in traditional herbal medicine systems across the Himalayan belt (India, Nepal, Bhutan, China) and has more recently gained interest for neuro-therapeutic applications. Ethnomedicinally, the rhizomes and roots of V. jatamansi have been used for insomnia, anxiety, epilepsy, digestive disorders and women's health. This species is often contrasted with the well-known European valerian, Valeriana officinalis L., which has been widely marketed for sleep disorders in the West (European Medicines Agency, 2011). Although both share comparable therapeutic indications, their phytochemical profiles differ meaningfully especially in the nature and abundance of valepotriates, sesquiterpenes and iridoids thus motivating independent scientific exploration of V. jatamansi.
The pharmacological importance of V. jatamansi lies at the intersection of traditional use (for CNS disorders) and emergent evidence from phytochemical and mechanistic studies. Interest in harnessing natural products in neurodegenerative disorders, anxiety and insomnia makes V. jatamansi a promising candidate. The objective of this review is to provide a comprehensive, up-to-date consolidation of the phytochemistry and pharmacological findings (especially neuro-pharmacological) of V. jatamansi, critically assess mechanisms of action, safety and standardisation issues, and identify future research directions.
2. Botanical Description and Distribution
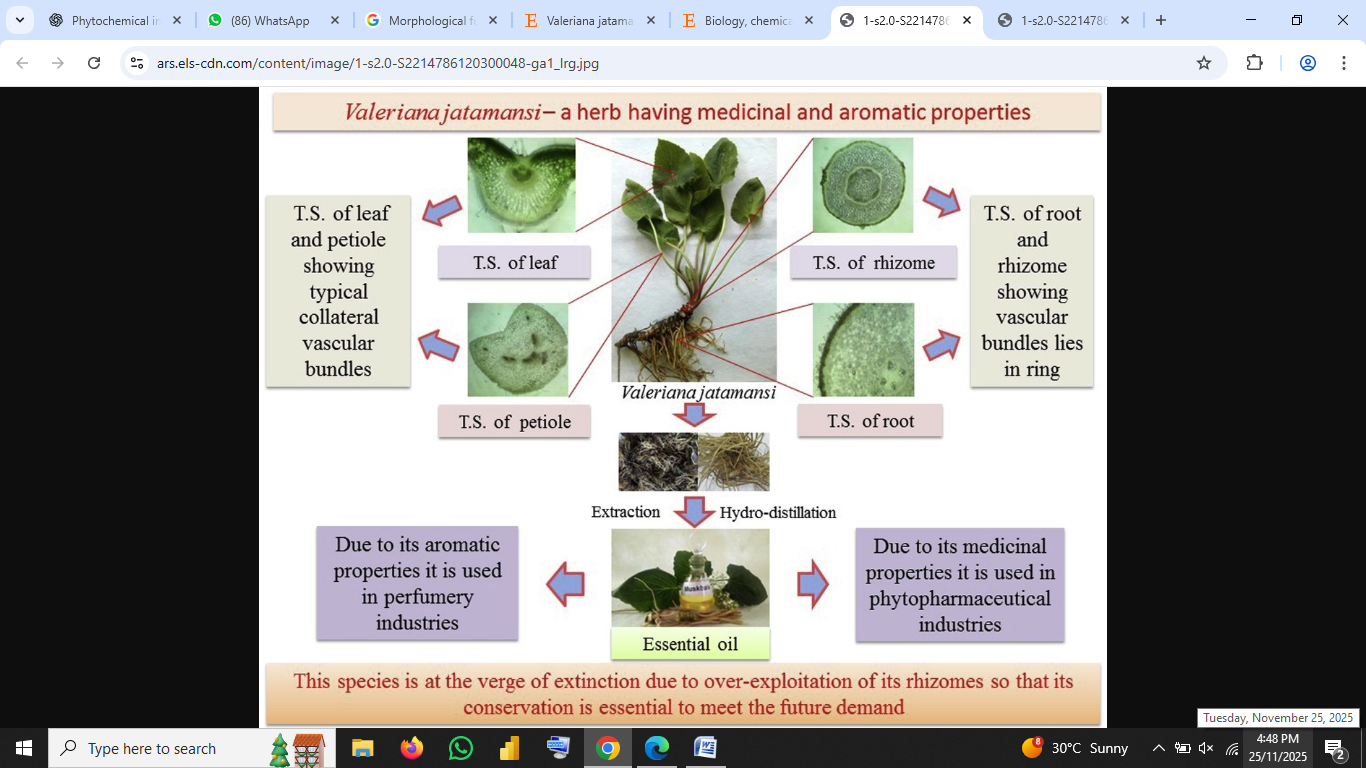
Fig no.1: Morphological features of Valeriana jatamansi Jones, showing root, rhizome, leaves, and inflorescence
Taxonomical classification
Morphological features
V. jatamansi is a small perennial herb, typically 20–70 cm in height, with fibrous rhizomes and thickened internodes. The stems are usually branched and densely pubescent. Basal leaves are cordate to ovate, 2–10 cm long × 1.5–8 cm wide, with 5–9 basal veins and pubescence on both surfaces. Cauline leaves are smaller and sessile. Inflorescences are terminal cymes; the small corollas (~6 mm) are white to slightly reddish, and the pistil and stamens protrude beyond the corolla tube (Ma et al., 2021 citing Jugran et al., 2013). Flowering occurs May–July; fruiting June–September.
Geographic distribution & ecology
The species is native to the Himalayan region (India: Uttarakhand, Himachal Pradesh; Nepal; Bhutan; China: Yunnan, Sichuan, Guizhou, Shanxi) and extends into parts of Pakistan and Myanmar (Ma et al., 2021; Li et al., 2022). It is typically found at altitudes of ~1000–3000 m above sea level, growing in grasslands, forest margins, streamsides, shady slopes and alpine meadows. Ecological and altitudinal variation affects its phytochemical profile (Jugran et al., 2013; Ma et al., 2021). For example, altitude has been shown to modulate valerenic acid, total phenolics and flavonoids in wild vs planted individuals (Jugran et al., 2016). Cultivation is being advanced to relieve wild-harvest pressure (Pandey et al., 2023).
Parts used and collection methods:
Traditionally, the rhizomes and roots (Radix et Rhizoma) are harvested, usually in late summer or autumn when secondary metabolite accumulation is optimal. After harvest, cleaning, shade-drying (or low-temperature oven drying to preserve key constituents) is advisable; for example, recent optimisation of drying conditions improved yield and stability of key iridoids (Xiao et al., 2025). In herbal commerce, standardised extracts or essential oils are prepared from roots/rhizomes; aerial parts are less commonly used for major bioactivity.
3. Ethnomedicinal and Traditional Uses
The medicinal use of V. jatamansi (Tagar) is entrenched in Ayurvedic, Unani and Tibetan medical texts, as well as Chinese ethnomedicine (Ma et al., 2021; Li et al., 2022). Traditional uses include:
The traditional breadth of indications underlines the herb’s potential for neuro-pharmacological and systemic therapeutic exploitation. The comparative use of V. officinalis in European herbalism underscores the rationale to examine V. jatamansi scientifically the present review aims to consolidate that effort.
4. Phytochemical Composition
The phytochemical profile of V. jatamansi is complex and multi-class; major constituent classes and key compounds are summarised in Table 1.
|
Compound name |
Chemical class |
Molecular formula |
Plant part isolated |
Reported pharmacological activity |
|
Valtrate |
Iridoid ester (valepotriate) |
C??H??O? |
Root/rhizome |
Anxiolytic, anticonvulsant (Yan et al., 2018) |
|
Didrovaltrate |
Iridoid ester |
C??H??O?? |
Root/rhizome |
Sedative/anticonvulsant |
|
Acevaltrate |
Iridoid ester |
C??H??O?? |
Root/rhizome |
CNS modulatory |
|
Jatamansone (-) |
Sesquiterpene (valeranone) |
C??H??O |
Rhizome essential oil |
Antioxidant, neuroprotective |
|
Patchoulol |
Sesquiterpene alcohol |
C??H??O |
Rhizome oil |
CNS effect |
|
β-caryophyllene |
Sesquiterpene |
C??H?? |
Rhizome/rhizome oil |
Anti-inflammatory |
|
Quercetin |
Flavonoid |
C??H??O? |
Whole plant |
Antioxidant, GABA modulator |
|
Apigenin |
Flavonoid |
C??H??O? |
Whole plant |
CNS calming |
|
Gallic acid |
Phenolic acid |
C?H?O? |
Roots/leaves |
Antioxidant, neuroprotective |
|
Actinidine |
Alkaloid |
C??H??N |
Rhizome |
Sedative/spasmolytic |
|
Valerine |
Alkaloid |
C?H??N |
Rhizome |
CNS modulation |
|
Bornyl acetate |
Monoterpene ester |
C??H??O? |
Essential oil of rhizome |
Aromatherapeutic, sedative |
|
Lignan (un-named) |
Lignan glycoside |
— |
Root/rhizome |
Cytotoxic/antitumour |
*Molecular formulas are illustrative.
Sources: Iridoids and sesquiterpenoids from V. jatamansi (Dong et al., 2015) — Fitoterapia; Li et al. 2022 review.
Fig no.2: Representative chemical structures of major constituents of Valeriana jatamansi: (A) Valtrate, (B) Didrovaltrate, (C) Jatamansone, (D) Quercetin, (E) Actinidine.
Extraction and analytical methods:
Extraction methods for V. jatamansi typically involve hydroalcoholic extraction of rhizome/roots, followed by fractionation (e.g., silica gel, Sephadex, resin D101) for valepotriates/iridoids. Essential oils are obtained by hydrodistillation and analysed via GC–MS (Verma et al., 2011; Bhatt et al., 2012). The valepotriates are unstable, prone to hydrolysis, hence require careful handling (mass spectrometric profiling studies: Zhang et al., 2013). Analytical techniques include HPLC, LC-MS, GC-MS, UPLC–Q-TOF and NMR for structural elucidation. For example, Chen et al. (2017) isolated new iridoids using Phytochemistry (2017;141:156–161). The review by Li et al. (2022) compiled 445 secondary metabolites across Valeriana spp., emphasising iridoids, sesquiterpenes and lignans.
Variations in chemical profile
Geographic origin, altitude, cultivation status (wild vs planted) and harvesting season affect the chemical makeup. Jugran et al. (2016) demonstrated altitude-linked variation of valerenic acid, total phenolics, flavonoids in V. jatamansi. Raina & Negi (2015) reported essential-oil composition differences in Himalayan populations. Drying method also influences constituent stability: optimisation of drying (sun vs shade vs oven) improved iridoid retention (Xiao et al., 2025). Cultivation efforts and in-vitro propagation have been proposed to conserve wild stocks and ensure quality (Cell Press Heliyon, 2023).
Overall, while valepotriates and flavonoids stand out as major constituent groups, sesquiterpenes and essential-oil components provide significant biologic activity. The later sections will correlate these phytochemicals with pharmacological endpoints.
5. Pharmacological Activities
This section delineates the pharmacological activities of V. jatamansi with mechanistic insights, organised by category.
5.1 CNS and Neuropharmacological Activities
A wealth of studies delineate the CNS-modulatory potential of V. jatamansi.
Sedative and anxiolytic: The iridoid-rich fraction from V. jatamansi exhibited anxiolytic activity at 6, 9 and 12 mg/kg in mice (elevated plus-maze, light-dark box, Vogel’s conflict) and was shown to regulate GABA levels in brain tissue (PMID 28782383). Additionally, a valepotriate (valtrate) study in rats indicated anxiolytic action with changes in serum corticosterone (Wang et al., 2014). Mechanistically, modulation of GABAergic neurotransmission (enhanced brain GABA, glycine) and involvement of the HPA axis have been indicated (Yan et al., 2010). The literature suggests parallels with the well-studied V. officinalis, yet V. jatamansi appears to possess distinct constituents (e.g., novel iridoids) contributing to nervous-system effects.
Anticonvulsant: Some older work reports anticonvulsant activity of valepotriates in tonic-clonic seizure models (Wu et al., 2017). The N-type calcium channel antagonism (Cav2.2) by specific iridoids isolated from V. jatamansi (EC?? ~3.3–4.8 μM) suggests a mechanistic basis for nociceptive-/convulsion-modulatory potential (Ma et al., 2021 — reviewing Dong et al., 2017). This ion-channel modulation is noteworthy for neurotherapeutic claims.
Neuroprotective: Iridoids from V. jatamansi conferred protection in PC12 and MPP?-induced dopaminergic neuronal death models (BBB 76:1401-1406) (2018). More recently, an iridoid-rich fraction (IRFV) promoted axonal regeneration and motor-functional recovery in a spinal-cord-injury (SCI) rat model via PI3K/Akt activation, increased BDNF and NGF expression (Front. Mol. Neurosci. 2024) (turn1search2). This suggests potential beyond symptomatic sedation, into regenerative neurology.
Mechanistic summary
Comparison with V. officinalis suggests that while both modulate GABA_A receptors, differing constituent spectra (e.g., more iridoids/valepotriates in V. jatamansi) may afford distinct pharmacologic profiles.
5.2 Antioxidant and Anti-inflammatory Activities
Oxidative stress and neuroinflammation are common drivers of neurodegenerative and neuropsychiatric disorders; accordingly, V. jatamansi has shown antioxidant and anti-inflammatory capacity.
These antioxidant–anti-inflammatory properties plausibly underpin the neuro- and hepatoprotective functions of the herb—serving as a plausible adjunct for oxidative-stress-driven CNS pathologies.
5.3 Antimicrobial and Antifungal Activities
Several studies describe antimicrobial potential of V. jatamansi. For example, references indicate antibacterial and antifungal activities of extracts and isolated sesquiterpenoids (Liu et al., 2017). While not central to neuro-therapeutic claims, these effects may support wound-healing and traditional uses in infections.
5.4 Anticancer Properties
Emerging evidence indicates cytotoxic effects of V. jatamansi constituents:
5.5 Cardioprotective and Hepatoprotective Effects
V. jatamansi extracts have been shown to modulate lipid profiles, reduce oxidative-injury markers and protect hepatic enzyme systems in rat models of experimental hyperlipidaemia (Chinese J Exp Trad Med Formulae 2012) (Ma et al., 2021). While mechanistic depth is limited, the data align with antioxidant/anti-inflammatory profiles.
5.6 Endocrine and Metabolic Effects
Studies indicate regulation of glucose metabolism and stress-hormone modulation: for example, iridoid-rich fraction reduced serum corticosterone, improved lipid profiles and regulated insulin/glucose in animal models (Zhu et al., 2016). Such adaptogenic or metabolic effects could complement CNS-therapeutic contexts (anxiety-linked metabolic dysregulation).
5.7 Other Activities
Other reported effects include adaptogenic/immunomodulatory (splenic lymphocytes/macrophage modulation in mice: Ma et al., 2010), spasmolytic/gastroprotective (ethoxyviburtinal-11 modulated colonic contractility: China J Trad Med Pharma 2016), wound-healing, and gastroenteric benefits in paediatric rotavirus enteritis (Ma et al., 2021). While less explored mechanistically, these broaden the therapeutic horizon.
Critical commentary:
While pharmacological studies are numerous, there are limitations: many investigations remain in vitro or animal-based, human clinical data are scarce, pharmacokinetic/pharmacodynamic (PK/PD) relationships are largely undefined, and standardisation of extracts remains inconsistent. Nonetheless, the cumulative evidence supports the neuro-therapeutic promise of V. jatamansi.
6. Mechanistic Insights in Neuropharmacology
Delving deeper into mechanistic underpinnings, several pathways merit emphasis:
6.1 GABAergic system modulation
Multiple studies indicate that V. jatamansi extracts and isolated compounds enhance brain GABA and glycine content and modulate GABA_A-receptor activity. An iridoid fraction increased GABA levels in rats and produced anxiolytic behaviour (PMID 28782383). In the context of the HPA-axis, local GABAergic influences on the HPA axis are well-documented (Cullinan & Ziegler, 2008), which may underpin stress-modulatory benefits. Although direct patch-clamp data on V. jatamansi constituents are somewhat limited, the presence of flavonoids such as 6-methylapigenin (also present in valerian species) suggests positive modulation of GABA_A receptors (Fernández et al., 2005). By enhancing inhibitory neurotransmission, V. jatamansi may exert sedative/anxiolytic effects analogous to those of benzodiazepine-type modulators but with distinct phytochemical profiles.
6.2 Ion channel modulation
Iridoids from V. jatamansi were shown to inhibit N-type voltage-gated calcium channels (Cav2.2) with EC?? values ~3–5 µM, providing a mechanistic basis for analgesic/anticonvulsant effects (Dong et al., 2017). The modulation of ion channels may work synergistically with GABAergic modulation to stabilise neuronal excitability, thereby contributing to anticonvulsant and neuroprotective actions.
6.3 Neurotrophic and regenerative signaling
Recent work (Wang et al., 2024) demonstrated that the iridoid-rich fraction (IRFV) promotes axonal regeneration and motor recovery in a spinal-cord-injury model via the PI3K/Akt pathway and up-regulation of neurotrophic factors BDNF and NGF. This is compelling because it places V. jatamansi in the realm of neuro-repair rather than only symptomatic relief. Activation of PI3K/Akt is well-established in neuronal survival and plasticity, so these findings open possibilities for neurodegenerative disease contexts (e.g., Alzheimer’s, Parkinson’s).
6.4 Monoamine/MAO modulation and serotonergic interactions
While specific data on V. jatamansi are limited, reviews of the genus indicate potential for MAO-A/B inhibition (Li et al., 2022). For example, modulation of 5-HT and 5-HIAA levels in IBS-model rats was observed with V. jatamansi extract (Yan et al., 2011). This suggests a multi-modal mechanism: GABAergic plus serotonergic/monoaminergic interactions, which combined may underlie anxiolytic/antidepressant effects.
6.5 Antioxidant/anti-inflammatory neuroprotection
Oxidative stress and neuroinflammation are upstream drivers of neurodegeneration and neuronal injury. The potent radical-scavenging, SOD/catalase activation and cytokine-modulation effects of V. jatamansi (Wang et al., 2017; Zhu et al., 2016) can thus support neuronal resilience. In concert with ion-channel and neurotrophic signalling modulation, these effects may afford broad neuroprotective potential.
6.6 Summary: Relevance for neurodegenerative diseases and anxiety disorders
The convergence of mechanisms — inhibitory neurotransmission (GABA), ion-channel stability, neurotrophic signalling (PI3K/Akt/BDNF/NGF), and antioxidant/anti-inflammatory effects positions V. jatamansi as a potential multi-target neurotherapeutic. In anxiety disorders and insomnia, the GABAergic/sedative axis is immediately relevant; in neurodegenerative disease (e.g., Alzheimer’s, Parkinson’s) or spinal-cord injury, the neuro-repair/regeneration axis becomes pertinent. Although clinical evidence is yet lacking, mechanistic plausibility is strong.
7. Toxicity, Safety, and Standardization
Toxicity and safety: Acute and sub-chronic toxicity assessments exist. For example, a sub-chronic toxicology study of iridoid-rich fraction (IRFV) in mice and rats (J Ethnopharmacol 2015;172:386-94) found no significant adverse effects at tested doses (Ma et al., 2021). These data indicate a favourable safety margin for experimental animal administration. However, human safety data (clinical trials) are absent or minimal.
Standardisation challenges:
Quality control and GACP: Good Agricultural and Collection Practices (GACP) are recommended given the endangered status of wild populations and the variability in cultivation. Cultivation under controlled conditions, propagation in vitro, and defined harvesting/drying protocols (Xiao et al., 2025) enhance consistency. Analytical methods (HPLC, LC–MS) can quantify major marker compounds (e.g., valtrate, jatamansone, quercetin) and ensure batch-to-batch uniformity.
In summary, while V. jatamansi appears relatively safe in animal studies, translation to human use demands formal toxicology, pharmacokinetics and standardised formulations.
8. Recent Advances and Future Prospects
Analytical and phytochemical advances (2020–2025):
Formulation progress:
Nanocarrier/microemulsion platforms for improved bioavailability of herbal extracts are beginning to appear in herbal-neuropharmacology domains (though specific reports for V. jatamansi are only emerging). Its incorporation into polyherbal neurotherapeutic formulations is plausible given its GABAergic, antioxidant and neurotrophic profile.
Research gaps and opportunities:
9.CONCLUSION
Valeriana jatamansi Jones emerges as a scientifically substantiated herb with considerable neuro-pharmacological promise. Its phytochemical richness spanning valepotriates, iridoids, sesquiterpenes, flavonoids, alkaloids and essential oils supports a pan-optic pharmacology: sedation/anxiolysis, neuroprotection/regeneration, antioxidant/anti-inflammatory, antimicrobial and anticancer effects. Mechanistic explorations demonstrate modulation of GABAergic neurotransmission, ion-channel stabilisation, neurotrophic signalling (PI3K/Akt, BDNF/NGF) and oxidative-stress defence. From a therapeutic perspective, the CNS-modulating and neuro-regenerative dimensions are especially compelling for anxiety disorders, insomnia, spinal-cord injury and potentially neurodegenerative disease.
However, to actualise its clinical potential, several hurdles remain: rigorous human clinical trials, detailed pharmacokinetic mapping, standardised extract development, and sustainable cultivation with chemical-profile consistency. Given the increasing global interest in herbal neurotherapeutics, V. jatamansi is well-positioned as a promising candidate in the botanical arsenal. The path forward calls for translational commitment: from bench to bedside, ensuring safety, efficacy and quality for neuro-therapeutic application.
REFERENCES

Tajas Gavade*, Sanika Nikam, Dhanraj Jadge, Phytochemical and Pharmacological Insights into Valeriana Jatamansi Jones (Tagar/Indian Valerian): A Promising Neuropharmacological and Therapeutic Herb, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 11, 4054-4066 https://doi.org/10.5281/zenodo.17719337
 10.5281/zenodo.17719337
10.5281/zenodo.17719337
