
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
1 California Institute of Technology, Pasadena, California 91126, USA.
1, 2,3,4,5,6,7 Cell and Protein Sciences, Aragen Life Sciences, Hyderabad, Telangana 500076, India.
PROteolysis Targeting Chimeras (PROTACs) are bifunctional molecules that promote targeted protein degradation by recruiting an E3 ubiquitin ligase to the protein of interest (POI), forming a ternary complex that facilitates ubiquitination and proteasomal degradation. The dynamic nature of these complexes presents challenges for characterization. Conventional methods such as fluorescence polarization (FP), time-resolved FRET (TR-FRET), and AlphaLISA lack real-time and label-free kinetic resolution, while isothermal titration calorimetry (ITC) demands large sample quantities and low throughput. Here, we utilize surface plasmon resonance (SPR) to characterize the binding affinity and cooperativity of a ternary complex comprising cyclin-dependent kinase 2 (CDK2), the PROTAC CPS2, and the CRBN–DDB1 E3 ligase complex. Using the Biacore T200 system, we measure real-time, label-free binary and ternary binding kinetics, revealing a cooperativity factor (?) of approximately 98, indicative of strong positive cooperativity. Our findings highlight SPR as a robust platform for quantitative assessment of PROTAC-induced ternary complexes.
PROTACs represent an innovative therapeutic strategy by promoting the formation of ternary complexes that facilitate ubiquitination and subsequent degradation of target proteins via the UPS [1,2]. This approach has redefined drug discovery, allowing therapeutic targeting of previously “undruggable” proteins. Central to this mechanism is the CRBN–DDB1 complex, a key component of the Cullin-RING E3 ubiquitin ligase system. CRBN acts as a molecular glue that recruits substrates for ubiquitination, while DDB1 stabilizes the complex through interaction with Cullin-4, ensuring substrate-specific degradation [3]. Understanding ternary complex dynamics is critical for optimizing PROTAC performance. However, conventional techniques such as FP, TR-FRET, and AlphaLISA depend on labeled components and are limited in throughput, making them less suitable for rapid PROTAC screening [4]. In contrast, SPR is a powerful, label-free technique capable of measuring real-time biomolecular interactions. By detecting changes in the refractive index at the sensor surface, SPR provides precise kinetic and affinity data for both small molecules and large complexes [5].
Despite progress in PROTAC development, there remains a lack of kinetic and cooperative binding data for ternary complexes involving CDK2, a kinase of clinical interest. Moreover, while SPR offers a powerful, label-free method for studying such interactions, its application in PROTAC evaluation—especially in the Indian research context—has been limited. PROTACs are increasingly being explored for targeting disease-relevant proteins across oncology, including cyclin-dependent kinases such as CDK2, which play pivotal roles in cancer cell proliferation and have been shown to be synthetically lethal in MYCN-overexpressing cancers [6,7]. Recent compounds like CPS2 have shown promise in acute myeloid leukemia (AML) models through targeted CDK2 degradation [8]. Efficient characterization of such degrader molecules at the preclinical stage is essential to enable downstream translational studies, making robust methods like SPR crucial for early-stage evaluation.
In this study, we have used SPR based binding assay to characterize the binding kinetics and cooperativity of a PROTAC-induced ternary complex involving CDK2 and CRBN–DDB1. Recombinant CDK2 and CRBN–DDB1 proteins were expressed and purified for binding studies to determine the respective dissociation constants (KD) and the cooperativity factor (α). Our findings support the use of SPR as a high-quality platform for validating and characterizing PROTAC-induced complex formation. To our knowledge, this is the first report from an Indian research facility to use SPR for quantifying PROTAC ternary complex cooperativity (α ≈ 98), with additional insights into the impact of mammalian expression systems on assay development and reproducibility.
MATERIALS AND METHODS:
Plasmid Generation
PCR amplified DDB1 and 8His-TEV-3xFlag-GS-3C-Avi-GS3-CRBN were subcloned into NheI and XhoI digested pcDNA3.4 to generate pcDNA3.4-DDB1 and pcDNA3.4-8His-TEV-3xFlag-GS-3C-Avi-GS3-CRBN constructs respectively. Similarly, GST-3C-CDK2-6His was PCR-amplified and subcloned into XbaI- and XhoI digested pET28a vector to generate pET28A- GST-3C-CDK2-6His.Plasmids pcDNA3.4-8xHis-TEV-3xFlag-3C-Avi-linker-CRBN and pcDNA3.4-DDB1 were purified using the Endotoxin-Free Quanta Maxi Kit (MDI, Cat# QXEK0025) and resuspended in endotoxin-free LAL reagent water (Charles River, Cat# W110).
Protein Expression and Purification
Expi293F™ cells (Thermo Fisher Scientific, Cat# A14528) were maintained in a shaker incubator at 37?°C with 8% CO2 and tested regularly for mycoplasma contamination using the Mycoplasma Detection Kit (ATCC, Cat# ATCC-30-1012K). Cells were passaged once they reached a density of 5 × 106 cells/mL and were not used beyond passage 30. CRBN and DDB1 plasmid constructs were transfected into Expi293FTM cells using the ExpiFectamineTM 293 Transfection Kit (Thermo Fisher Scientific, Cat# A14524) to express the CRBN-DDB1 protein complex. CRBN-DDB1 complex was purified by affinity chromatography followed by ion exchange using HiTrap Capto™ Q anion exchange column. The purity of the complex was further polished by size exclusion chromatography leading to the purified CRBN-DDB1 complex. The purity of the protein complex was verified on aSE-HPLC and mass integrity was confirmed by intact mass analysis using mass spectrometry. CDK2-His was expressed in BL21 Star™ (DE3) E. coli strain (as a GST-3C fusion and purified by affinity chromatography. The GST tag was removed using 3C protease. The purified recombinant proteins were used in SPR-based binding assays.
Surface Plasmon Resonance (SPR) Binding Assay
SPR experiments were performed on a Biacore T200 instrument using Biacore T200 Control and Evaluation Software v3.4 (Cytiva). CDK2 was immobilized at 20 °C onto a Series S NTA chip (Cytiva; Cat# BR100532) in 1X PBS-P+ buffer (Cytiva; Cat# 28995084). The surface was charged with Ni2+ (Cytiva; Cat# 28995043) after conditioning with 350 mM EDTA. CDK2 was then captured to a density of 2500–3000 response units (RU) using EDC/NHS coupling. Unreacted sites were blocked with ethanolamine. CRBN-DDB1 was immobilized on a Series S SA chip (Cytiva; Cat# BR100531) via biotin-streptavidin interaction. The chip was pre-conditioned with 50 mM NaOH/1 M NaCl. Biotinylated CRBN-DDB1 (100 µg/mL) was then injected to achieve 4500–5500 RU. Unoccupied sites were blocked using biocytin. For both CDK2 and CRBN-DDB1, the reference cell was prepared identically, except that no protein was injected, leaving it empty to eliminate any non-specific binding to the chip surface. For binary interactions study, CPS2 (MedChemExpress, HY-141680) was serially diluted (0.0027–2 µM) and injected on to immobilized CDK2 or CRBN-DDB1 in multi-cycle kinetics format (association: 120 s, dissociation: 400 s, flow rate: 30 µL/min). Duplicate injections and blanks were included, with syringe washes of 50% DMSO in between cycles. For all kinetics experiments, 1X PBS-P+ was supplemented with 2% DMSO for solubility of CPS2. For evaluation of ternary complex confirmation, 1:1 mixture of 2 µM CPS2 (in 4% DMSO) and 20 µM CDK2 was prepared, yielding a final concentration of 1 µM CPS2 and 10 µM CDK2 in 2% DMSO. Serial dilutions were made to this mix, CDK2 and CPS2, to generate a 5-point dilution series (1 µM to 0.0039 µM CPS2; 10 µM to 0.502 µM CDK2). Injections were performed using single-cycle kinetics format (association: 100 s, dissociation: 720 s, flow rate: 100 µL/min).
The data obtained from SPR studies were double-referenced using the reference cell data and the data obtained from blank runs (0 µM CPS2) in Biacore Evaluation Software. The affinity KD for CDK2:CPS2 and CRBN-DDB1:CPS2 binary binding was determined using a 1:1 Langmuir binding model (steady state) fitting. For ternary complex confirmation assay, an apparent affinity KD was determined using the same method as described previously for binary binding. The cooperativity factor (α) was estimated for evaluating the stability of the ternary complex using the following equation:
α = KDBinary / KDTernary
RESULTS
To evaluate the stability of ternary complexes formed through PROTAC-mediated interactions between E3 ligases and target proteins, recombinant purification was performed for CDK2 (the target protein) and CRBN-DDB1 (a key component of the CUL4-RING E3 ubiquitin ligase). Recombinant CDK2 was expressed in the E. coli expression system and was purified to homogeneity confirmed using SDS-PAGE, analytical size exclusion HPLC and intact mass analysis using mass spectrometry (Figure 1A). Recombinant CRBN and DDB1 were co-expressed in the mammalian expression system and were purified as a complex confirmed using SDS-PAGE, analytical size exclusion HPLC and intact mass analysis using mass spectrometry (Figure 1B).
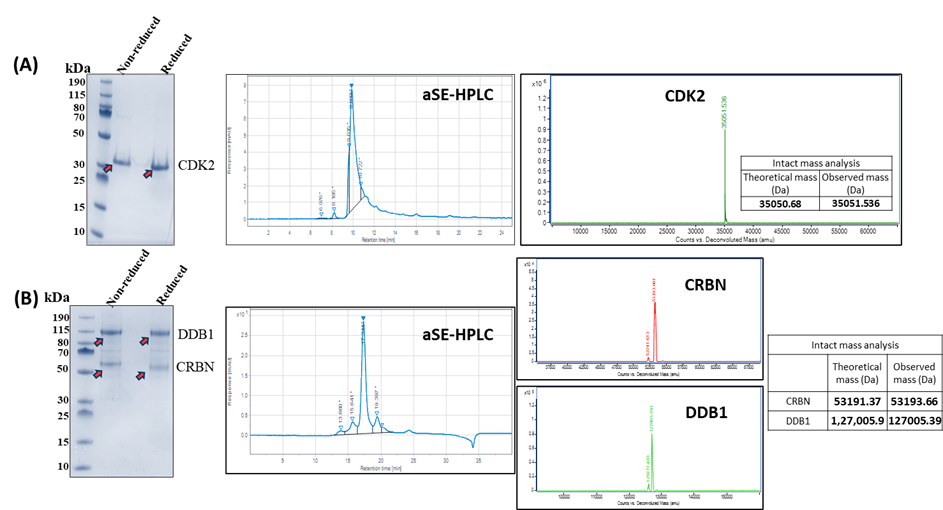
Figure 1. Recombinant purified (a) CDK2 and (b) CRBN-DDB1 complex analysed by SDS-PAGE, analytical size exclusion HPLC, and mass spectrometry to confirm purity and molecular weight integrity.
Legend: SDS-PAGE: Sodium dodecyl sulfate polyacrylamide gel electrophoresis used to evaluate protein purity. HPLC: High-performance liquid chromatography performed for size exclusion analysis. Mass spectrometry confirmed the molecular weight integrity of purified proteins. CDK2: Cyclin-dependent kinase 2; CRBN: Cereblon; DDB1: DNA damage-binding protein 1.
CPS2 is a first-in-class, highly potent, selective, and irreversible PROTAC that degrades CDK2, with an IC?? of 24 nM [8]. CPS2 has been shown to degrade CDK2 across multiple Accute myeloid leukemia cell lines, including NB4, U937, OCI-AML2, OCI-AML3, and Kasumi-1and hence is used in AML associated research and development [9]. However, there is no report in existing literature depicting the direct binding of CPS2 to CDK2 and CRBN-DDB1 complex using a biophysical method. For this reason, in this study we use SPR to study PROTAC binding, including ternary complex formation and affinity measurements. Prior to the ternary complex study, the binary interactions of CPS2 with CDK2 and CRBN-DDB1 were individually characterized to estimate their binding affinities. These measurements informed the selection of appropriate concentration ranges for ternary complex evaluation and enabled assessment of ternary complex stability through the cooperativity factor.
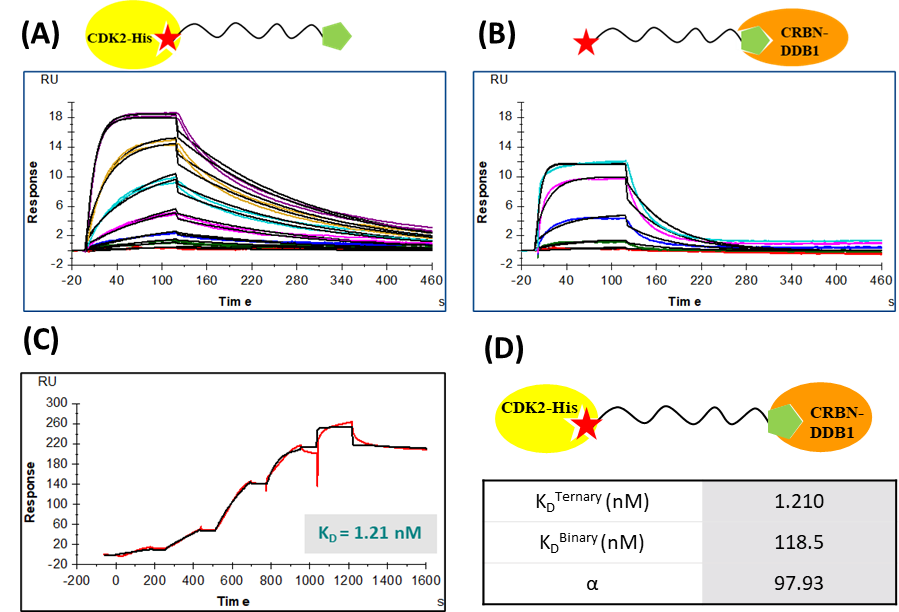
Figure 2: SPR binding assay. (a) Multicycle binding kinetics of CPS2 binding to CDK2-His. (b) multicycle binding kinetics of CPS2 binding to CRBN-DDB1 complex. (c) Binding kinetics of CRBN-DDB1:CPS2:CDK2-His ternary complex. (d) Cooperativity factor, α, estimation.
Legend: SPR: Surface plasmon resonance used to measure binding interactions. CPS2: Small molecule ligand under study. His: Histidine tag used for protein purification. Data represent mean binding kinetics obtained from multiple cycles to assess affinity and cooperativity. Binary binding assay between CDK2 and CPS2 exhibited a dissociation constant (KD) of 136 .4 nM, depicting a well fitted kinetic profile through sensorgrams displayed in Figure 2A. An estimated KD of 136.4 nM indicates specific binding affinity, consistent with stable engagement of the PROTAC with its target protein CDK2. Figure 2B shows precisely fitted sensorgrams displaying the binary interaction between CRBN-DDB1 and CPS2 with a slightly tighter binding profile with a KD of 118.5 nM. Together, these results confirm that CPS2 can independently bind both the target and E3 ligase CRBN-DDB1 and CPS2 demonstrating effective interaction of CPS2 with both the components. The binding response curves obtained for each interaction from the binding assay, plotted as response units (RU) against PROTAC concentration (nM), illustrate distinct kinetic profiles and relative binding strengths. Both binary interactions exhibit comparable nanomolar affinities, which should help facilitate the formation of a ternary complex between the PROTAC, CDK2, and CRBN-DDB1.
The PROTAC mediated ternary complex formation was studied in SPR using the single cycle kinetics format to capture the ternary complex and avoid the dissociation of the complex during the assay. For the ternary complex study, CRBN-DDB1 was immobilized on the chip due to its higher molecular weight compared to CDK2 and also, its stock concentration was insufficient for titration onto a CDK2-coated surface. A mixture of CPS2 and CDK2 was titrated onto the CRBN-DDB1 complex and was prepared with CDK2 at 10X of the CPS2 concentration, ensuring that during titration onto the CRBN-DDB1 surface, all CPS2 molecules are saturated with CDK2 and only the ternary complex is captured during kinetics. SPR sensorgrams displayed well-fitted kinetic profiles and reproducible data, supporting the specificity of the interactions.
CDK2:CPS2 titration on CRBN-DDB1 surface yielded a significantly higher binding affinity, with an apparent affinity KD of 1.21 nM (Figure 2C). This elevated affinity indicates that the CDK2:CPS2 complex binds more strongly to CRBN-DDB1 than the individual binary interactions, suggesting enhanced stability of the ternary complex. To further confirm the stability of ternary complex interaction, cooperativity factor (α) was estimated as the ratio of binary (118 nM) and ternary (1.21 nM) KD values (Figure 2D). An α of ~98 was computed indicating strong positive cooperativity, suggesting an almost 100-fold increased stabilization of the ternary complex upon simultaneous interaction of CPS2 with CDK2 and CRBN-DDB1. The enhanced stability observed for the ternary complex further validates the ability of CPS2 to bridge CDK2 and CRBN-DDB1 effectively and underscores the cooperative nature of ternary complex formation.
It is most striking to see this elevated cooperativity factor, where the ternary complex CRBN-DDB1:CPS2:CDK2 demonstrates a substantially strengthened binding affinity. Cooperativity factor estimations have been done for other ternary complexes stability analysis too but this high cooperativity has not been observed for any other PROTAC-based ternary complex. This PROTAC-mediated interaction highlights the effectiveness of CPS2 as a glue, facilitating the association between the target protein CDK2 and the E3 ligase component CRBN-DDB1.
These results not only validate the PROTAC mechanism of action for CPS2 but also highlight the utility of SPR as a sensitive and reliable method for quantifying both binary and cooperative ternary binding events. The data in Figure 2 thus provide robust experimental support for positive cooperativity and efficient ternary complex stabilization. Together, these findings provide compelling biophysical evidence for CPS2-mediated ternary complex formation with strong positive cooperativity, highlighting the ability of SPR to resolve subtle differences in binding affinity and interaction dynamics. Having established the molecular basis of these interactions and the impact of protein expression systems on assay performance, we next discuss the broader implications of our results for PROTAC characterization and assay design.
DISCUSSION:
In this study, we successfully purified the target protein CDK2 and the E3 ligase complex CRBN-DDB1 to homogeneity, enabling detailed biophysical characterization of their interactions with a PROTAC molecule. The purification protocols were optimized to yield high-quality proteins suitable for SPR analysis, with both CDK2 and CRBN-DDB1 retaining their functional integrity post-purification, as confirmed by SDS-PAGE, size-exclusion chromatography and mass spectrometry analysis as shown in Figure 1. CRBN-DDB1 was originally purified to homogeneity from insect expression systems (Data not shown). However, initial SPR trials using CRBN-DDB1 expressed in insect cells showed surface dissociation after immobilization and during kinetics due to which sensorgrams observed were not appropriate. Transitioning to mammalian cells markedly enhanced chip immobilization and improved data reproducibility, likely due to increased complex stability. Hence, all SPR experiments were performed using the CRBN-DDB1 complex expressed in the mammalian expression system. This underscores the importance of protein source and expression system in SPR assay reliability and quality [10].
SPR was employed to investigate both binary and ternary interactions involving the PROTAC, CDK2, and CRBN-DDB1. Initial binary interaction studies revealed nanomolar binding affinities between CPS2 and each of its individual partners, CDK2 and CRBN-DDB1. These comparable affinities provided a strong foundation for ternary complex formation, suggesting that CPS2 could effectively bring together the two proteins. The binding affinity for the ternary complex was significantly higher (KD: 1.21 nM) than for binary complexes. This resulted in a cooperativity factor (α) of ~98, suggesting synergistic interactions that greatly stabilize the ternary complex. Such strong positive cooperativity is a key design goal in PROTAC development, as it often correlates with improved degradation efficacy, enhanced selectivity, and favourable pharmacodynamics [11]. Previous studies have also demonstrated that dissociation kinetics of ternary complexes influence degradation efficacy [12].
Overall, our findings underscore the potential of CPS2 as a robust molecular glue in PROTAC design, capable of mediating efficient and stable ternary complex formation between CDK2 and CRBN-DDB1. These insights contribute to the growing understanding of PROTAC-mediated degradation mechanisms and provide a framework for future optimization of targeted protein degradation strategies. These findings not only advance our understanding of PROTAC-mediated degradation of CDK2 but also demonstrate the feasibility and value of implementing high-resolution SPR assays in Indian research settings for mechanistic evaluation. A limitation of this study is that functional degradation was not evaluated in a cellular system; future experiments should assess whether the high cooperativity observed in biophysical assay translates into effective target degradation in live cell systems. Additionally, the inclusion of mutant or control PROTACs could help dissect the contribution of individual domains to binding kinetics and cooperativity.
CONCLUSION:
SPR offers a powerful and sensitive platform for the quantitative analysis of PROTAC-mediated ternary complexes. By enabling real-time measurement of molecular interactions, SPR allows detailed characterization of binding kinetics and cooperativity between the PROTAC, target protein, and E3 ligase components. In our study, the PROTAC candidate CPS2 exhibited strong positive cooperativity in forming a ternary complex with CDK2 and the CRBN-DDB1 E3 ligase complex. This cooperative binding behaviour not only confirms the ability of CPS2 to efficiently bring these proteins into close proximity but also validates its potential as an effective and selective degrader of CDK2. Our approach, leveraging SPR’s sensitivity and quantitative capabilities, provides a robust framework for characterizing PROTAC-induced ternary complexes, thereby supporting the rational optimization of PROTAC molecules during early-stage drug development. This methodology accelerates the identification of candidates with improved degradation efficiency and selectivity, ultimately enhancing the drug discovery pipeline.
ACKNOWLEDGMENTS:
We thank Aragen Life Sciences for supporting this research.
COMPETING INTERESTS:
The authors have no competing interests to disclose.
REFERENCES

Ridah Shaista Shanavas, Dhriti Kundu, Mohammed Khalid Soudagar, Vamsi Krishna Sanga, Shubhi Jha, Amit Mahendra Karole, Richa Arya, Surface Plasmon Resonance-Based Characterization of PROTAC-Induced Ternary Complexes Involving CDK2 and CRBN-DDB1, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 7, 2454-2461. https://doi.org/10.5281/zenodo.16029940
 10.5281/zenodo.16029940
10.5281/zenodo.16029940