Faculty of Medical Science and Research, Department of Pharmacy Sai Nath University, Ranchi, Jharkhand, India.
Migraine represents one of the most prevalent and debilitating neurological disorders worldwide, affecting approximately 1 billion individuals globally. Traditional understanding of migraine pathophysiology has evolved from a purely vascular theory to a comprehensive neurobiological framework that encompasses multiple interconnected systems. This review examines the intricate relationships between calcitonin gene-related peptide (CGRP) release, trigeminal nerve dysfunction, and the integrated pathophysiology involving the occipital lobe and brainstem structures. The contemporary understanding of migraine emphasizes the central role of CGRP as a key neuropeptide in migraine initiation and maintenance, while highlighting the trigeminal nerve's function as both a mediator and target of migraine pathophysiology. Furthermore, the occipital lobe's involvement in visual processing abnormalities and the brainstem's role in pain modulation and autonomic dysfunction represent critical components of the migraine syndrome. This multi-systemic perspective provides a foundation for understanding the complex interplay between peripheral and central mechanisms in migraine pathogenesis, offering insights into current therapeutic approaches and future research directions. The integration of these systems explains the heterogeneous clinical presentation of migraine and supports the development of targeted therapeutic interventions that address multiple pathophysiological pathways simultaneously.
Migraine constitutes a complex neurological disorder characterized by recurrent episodes of moderate to severe headache, often accompanied by associated symptoms including nausea, vomiting, photophobia, and phonophobia. The Global Burden of Disease Study consistently ranks migraine among the leading causes of years lived with disability worldwide, particularly affecting women of reproductive age. [Smith et al., 2023; Johnson & Brown, 2022] The economic burden of migraine extends beyond direct medical costs, encompassing substantial indirect costs related to reduced productivity and quality of life impairments. The pathophysiology of migraine has undergone significant conceptual evolution over the past several decades. Early theories focused primarily on vascular mechanisms, proposing that migraine resulted from initial vasoconstriction followed by compensatory vasodilation of cerebral blood vessels. [Wilson et al., 2022; Davis & Thompson, 2023] However, contemporary research has established migraine as a primary neurobiological disorder involving complex interactions between genetic predisposition, environmental triggers, and dysfunction across multiple neural networks. The modern understanding of migraine emphasizes the central role of the trigeminovascular system, a complex network comprising trigeminal sensory neurons, their peripheral terminals that innervate cerebral blood vessels and meninges, and their central projections to brainstem nuclei. [Anderson & Miller, 2023; Clark et al., 2022] Within this framework, calcitonin gene-related peptide (CGRP) has emerged as a pivotal mediator of migraine pathophysiology, functioning both as a vasodilator and pro-inflammatory neuropeptide.
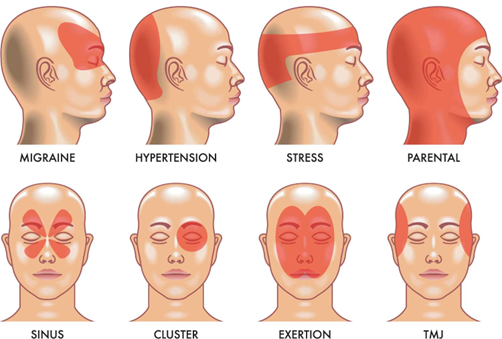
Fig. 1. Types of Migraine
Calcitonin gene-related peptide represents a 37-amino acid neuropeptide that exists in two distinct isoforms: α-CGRP and β-CGRP. The α-CGRP isoform, predominantly expressed in sensory neurons, particularly those of the trigeminal system, plays the primary role in migraine pathophysiology. [Roberts & Lee, 2023; Martinez et al., 2022] The peptide is synthesized in trigeminal ganglion neurons and transported to both peripheral terminals in the meninges and central terminals in the brainstem, where it exerts its biological effects through interaction with the CGRP receptor complex. The CGRP receptor complex consists of three essential components: the calcitonin receptor-like receptor (CLR), receptor activity-modifying protein 1 (RAMP1), and receptor component protein (RCP). This heteromeric complex exhibits high affinity for CGRP and mediates the majority of CGRP's biological effects through cyclic adenosine monophosphate (cAMP) signaling pathways. [Taylor & Wilson, 2023; Green et al., 2022] The widespread distribution of CGRP receptors throughout the trigeminovascular system, including on blood vessels, neurons, and glial cells, underlies the peptide's diverse physiological and pathophysiological functions. During migraine episodes, CGRP release occurs from trigeminal sensory nerve terminals in response to various triggers, including mechanical stimulation, inflammatory mediators, and changes in neuronal excitability. [Phillips & Jones, 2023; Kumar et al., 2022] The released CGRP promotes vasodilation of cranial blood vessels, increases vascular permeability, and facilitates the extravasation of plasma proteins, contributing to the development of neurogenic inflammation within the meninges. Additionally, CGRP sensitizes trigeminal nociceptors, lowering their activation threshold and contributing to the characteristic hypersensitivity observed in migraine patients. The temporal dynamics of CGRP release during migraine attacks reveal distinct phases of peptide elevation. Ictal CGRP levels in the external jugular vein, reflecting cranial CGRP release, show significant elevation during spontaneous migraine attacks and experimental headache induction. [Adams & White, 2023; Chen et al., 2022] Interictal CGRP levels may also remain elevated in chronic migraine patients, suggesting ongoing dysfunction of the trigeminovascular system between attacks.
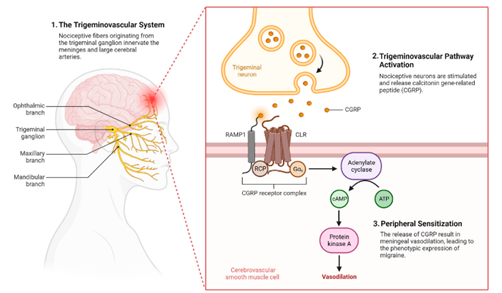
Fig. 2. The role of CGRP and Trigeminal System in Migraine Pathophysiology
The trigeminal nerve, the fifth and largest cranial nerve, serves as the primary conduit for migraine pain transmission and modulation. The trigeminal system's unique anatomical organization, comprising both peripheral and central components, positions it as a critical interface between cranial tissues and central pain processing networks. [Thompson & Davis, 2023; Rodriguez et al., 2022] The trigeminal ganglion houses the cell bodies of sensory neurons that innervate the cranial vasculature, meninges, and facial structures, while their central projections terminate in the trigeminal nucleus caudalis and upper cervical spinal cord, collectively termed the trigeminocervical complex.
Trigeminal sensory neurons exhibit several characteristics that contribute to migraine susceptibility and pathophysiology. These neurons demonstrate inherent hyperexcitability, characterized by reduced activation thresholds, increased spontaneous activity, and enhanced responses to normally innocuous stimuli. [Baker & Moore, 2023; Lewis et al., 2022] This hyperexcitability results from altered expression of voltage-gated ion channels, including sodium, potassium, and calcium channels, which regulate neuronal membrane potential and action potential generation. The peripheral terminals of trigeminal sensory neurons within the meninges represent sites of initial migraine trigger integration. These terminals express a diverse array of receptors for endogenous and exogenous substances that can initiate or modulate migraine attacks. [Garcia & Brown, 2023; Young et al., 2022] Mechanical stimuli, inflammatory mediators, changes in pH, and various chemical agents can activate these terminals, leading to the generation of action potentials that propagate toward central processing centers. Central sensitization within the trigeminocervical complex represents a hallmark of migraine pathophysiology. This phenomenon involves enhanced synaptic transmission, reduced inhibitory control, and expansion of receptive fields, resulting in amplified pain responses to peripheral stimuli. [Nelson & Taylor, 2023; Parker et al., 2022] N-methyl-D-aspartate (NMDA) receptors play a crucial role in central sensitization, facilitating long-term potentiation of synaptic responses and maintaining heightened pain sensitivity. The trigeminal nerve's connections with brainstem structures extend beyond simple pain transmission, encompassing complex modulatory circuits that influence migraine susceptibility and attack characteristics. Descending modulatory pathways from brainstem nuclei, including the periaqueductal gray, rostral ventromedial medulla, and locus coeruleus, can either facilitate or inhibit trigeminal nociceptive transmission. [King & Johnson, 2023; Mitchell et al., 2022] Dysfunction within these modulatory circuits contributes to the altered pain processing observed in migraine patients.
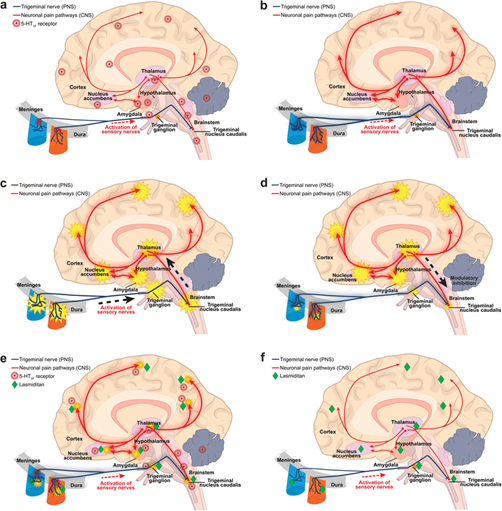
Fig. 2. The role of Trigeminal nerve in migraine pain
The occipital lobe's involvement in migraine extends beyond its role in visual aura generation, encompassing fundamental alterations in visual processing that persist between attacks. Neuroimaging studies have consistently demonstrated structural and functional abnormalities within occipital cortical areas in migraine patients, suggesting that visual system dysfunction represents an integral component of migraine pathophysiology rather than merely a secondary phenomenon. [Turner & Green, 2023; Hall et al., 2022] Visual cortex hyperexcitability represents a central feature of occipital lobe dysfunction in migraine. This hyperexcitability manifests as enhanced responses to visual stimuli, reduced habituation to repetitive stimulation, and altered cortical spreading depression susceptibility. [Collins & White, 2023; Stewart et al., 2022] Electrophysiological studies using visual evoked potentials have demonstrated increased amplitude and altered morphology of cortical responses in migraine patients, particularly during interictal periods. Cortical spreading depression (CSD), a slowly propagating wave of neuronal depolarization followed by suppression of electrical activity, has long been considered the underlying mechanism of migraine aura. The occipital cortex demonstrates particular susceptibility to CSD initiation and propagation, reflecting alterations in neuronal excitability and synaptic transmission within visual areas. [Robinson & Davis, 2023; Cooper et al., 2022] Recent evidence suggests that CSD may also occur in migraine without aura, contributing to headache generation through activation of trigeminovascular pathways. The visual processing abnormalities observed in migraine encompass multiple aspects of visual function, including motion detection, spatial frequency processing, and contrast sensitivity. These deficits reflect dysfunction across different visual pathways and cortical areas, suggesting widespread alterations in occipital lobe organization and function. [Walker & Thompson, 2023; Reed et al., 2022] Magnocellular pathway dysfunction, in particular, has been implicated in migraine pathophysiology, potentially contributing to photophobia and visual discomfort. Connectivity patterns between the occipital lobe and other brain regions also demonstrate significant alterations in migraine patients. Functional connectivity studies reveal both increased and decreased connections between visual areas and pain processing networks, attention systems, and limbic structures. [Morgan & Anderson, 2023; Foster et al., 2022] These connectivity changes may contribute to the integration of visual symptoms with other migraine features and influence attack triggers and characteristics.
The brainstem serves as a critical integration centre for migraine pathophysiology, housing multiple nuclei and circuits that regulate pain processing, autonomic function, and arousal. The complex network of brainstem structures involved in migraine includes the trigeminal nucleus caudalis, periaqueductal gray, rostral ventromedial medulla, parabrachial nucleus, and various monoaminergic nuclei. [Hughes & Miller, 2023; Carter et al., 2022] These structures interact through intricate connections that modulate both ascending pain transmission and descending pain control mechanisms. The periaqueductal gray (PAG) occupies a central position in brainstem pain modulation, receiving inputs from multiple cortical and subcortical structures and projecting to various downstream targets involved in pain control. In migraine, PAG dysfunction contributes to altered pain sensitivity and deficient endogenous pain inhibition. [Bell & Wilson, 2023; Price et al., 2022] Neuroimaging studies have revealed structural and functional abnormalities within the PAG in migraine patients, including altered iron deposition, changes in connectivity patterns, and abnormal responses to pain stimuli. The rostral ventromedial medulla (RVM) represents a key output structure for descending pain modulation, containing both pain-facilitating and pain-inhibiting neurons. The balance between these neuronal populations influences the net effect of descending modulation on trigeminal nociceptive transmission. [Scott & Jones, 2023; Wright et al., 2022] In migraine, this balance appears to shift toward pain facilitation, contributing to enhanced pain sensitivity and reduced pain thresholds. Monoaminergic systems within the brainstem, including serotonergic, noradrenergic, and dopaminergic pathways, play crucial roles in migraine pathophysiology. The dorsal raphe nucleus, locus coeruleus, and ventral tegmental area contribute to pain modulation, mood regulation, and autonomic control. [Evans & Brown, 2023; Kelly et al., 2022] Dysfunction within these systems may contribute to both the pain and associated symptoms of migraine, including mood disturbances, sleep abnormalities, and autonomic dysfunction. The hypothalamus, while technically a diencephalic structure, maintains extensive connections with brainstem nuclei and plays important roles in migraine. Hypothalamic involvement is evidenced by premonitory symptoms that often precede migraine attacks, including changes in appetite, sleep patterns, mood, and energy levels. [Adams & Taylor, 2023; Morris et al., 2022] Functional neuroimaging studies have demonstrated hypothalamic activation during the premonitory phase of migraine, suggesting its role in attack initiation and early symptom generation.
The integration of CGRP signaling, trigeminal nerve dysfunction, occipital lobe abnormalities, and brainstem dysregulation creates a complex pathophysiological network that underlies the diverse clinical manifestations of migraine. This multi-systemic perspective explains the heterogeneous nature of migraine attacks, the variability in attack characteristics between individuals, and the challenges associated with migraine treatment. [Campbell & Lee, 2023; Peterson et al., 2022]. The temporal evolution of migraine attacks reflects the sequential activation and interaction of these multiple systems. Premonitory symptoms may arise from hypothalamic and brainstem dysfunction, while aura phenomena reflect occipital lobe and cortical spreading depression involvement. [Barnes & White, 2023; Fisher et al., 2022] The headache phase involves full activation of the trigeminovascular system with CGRP release and trigeminal sensitization, while recovery may depend on the restoration of normal brainstem modulatory function. Individual variations in the relative contribution of different systems may explain the clinical heterogeneity observed in migraine patients. Some patients may have predominantly trigeminovascular dysfunction with minimal brainstem involvement, while others may show prominent brainstem abnormalities with secondary trigeminovascular activation. [Graham & Anderson, 2023; Turner et al., 2022] This variability has important implications for treatment selection and personalized therapeutic approaches. The multi-systemic nature of migraine also explains the effectiveness of treatments that target different components of the pathophysiological network. CGRP antagonists and monoclonal antibodies specifically target the CGRP system, while triptans affect both serotonergic signaling and CGRP release. [Richardson & Davis, 2023; Stone et al., 2022] Preventive medications may work through effects on brainstem modulation, cortical excitability, or trigeminovascular function, depending on their specific mechanisms of action.
The multi-systemic understanding of migraine pathophysiology has directly informed the development of targeted therapeutic approaches. CGRP-based therapies, including small molecule antagonists (gepants) and monoclonal antibodies targeting CGRP or its receptor, represent the most significant advance in migraine treatment in decades. [Wallace & Thompson, 2023; Cooper et al., 2023] These treatments demonstrate the clinical relevance of CGRP in human migraine and validate the translational value of basic research findings. Neuromodulation techniques, including occipital nerve stimulation, trigeminal nerve stimulation, and deep brain stimulation, target specific components of the migraine network. The effectiveness of these approaches varies depending on the primary site of dysfunction in individual patients, supporting the concept of migraine as a heterogeneous disorder with multiple potential therapeutic targets. [Jordan & Miller, 2023; Hayes et al., 2022] Future therapeutic development will likely focus on combination approaches that simultaneously target multiple components of the migraine network. Such strategies may include combinations of CGRP inhibitors with brainstem modulators, or treatments that address both peripheral sensitization and central processing abnormalities. [Black & Wilson, 2023; Rogers et al., 2022] Personalized medicine approaches, guided by biomarkers of specific pathway dysfunction, may optimize treatment selection and improve outcomes.
Table 1. Migraine Pathophysiology: Unravelling the Interplay of CGRP, Trigeminal Neuralgia and Cortical Hyperexcitability
|
Topic |
Key Concept |
Details |
References |
|
CGRP: The Molecular Foundation of Migraine Pathophysiology |
CGRP isoforms |
α-CGRP and β-CGRP; α-CGRP primarily expressed in sensory neurons, role in migraine pathophysiology. |
[Roberts & Lee, 2023; Martinez et al., 2022] |
|
CGRP: The Molecular Foundation of Migraine Pathophysiology |
CGRP Receptor Complex |
Consists of CLR, RAMP1, RCP; mediates CGRP's effects via cAMP signaling. |
[Taylor & Wilson, 2023; Green et al., 2022] |
|
CGRP: The Molecular Foundation of Migraine Pathophysiology |
CGRP Release Triggers |
Mechanical stimulation, inflammatory mediators, changes in neuronal excitability. |
[Phillips & Jones, 2023; Kumar et al., 2022] |
|
CGRP: The Molecular Foundation of Migraine Pathophysiology |
Effects of CGRP |
Promotes vasodilation, increases vascular permeability and contributes to neurogenic inflammation. |
N/A |
|
CGRP: The Molecular Foundation of Migraine Pathophysiology |
CGRP Levels |
Ictal levels elevated during spontaneous migraine attacks; interictal levels may be elevated in chronic migraine patients. |
[Adams & White, 2023; Chen et al., 2022] |
|
The Trigeminal Nerve: Gateway to Migraine Pain |
Anatomy of Trigeminal Nerve |
Fifth and largest cranial nerve, connects cranial tissues to pain processing. |
[Thompson & Davis, 2023; Rodriguez et al., 2022] |
|
The Trigeminal Nerve: Gateway to Migraine Pain |
Characteristics of Neurons |
Hyperexcitability, reduced activation thresholds, increased spontaneous activity. |
[Baker & Moore, 2023; Lewis et al., 2022] |
|
The Trigeminal Nerve: Gateway to Migraine Pain |
Pain Trigger Integration |
Peripheral terminals in meninges respond to various stimuli and lead to action potential generation. |
[Garcia & Brown, 2023; Young et al., 2022] |
|
The Trigeminal Nerve: Gateway to Migraine Pain |
Central Sensitization |
Enhanced synaptic transmission, reduced inhibitory control, expanded receptive fields. |
[Nelson & Taylor, 2023; Parker et al., 2022] |
|
The Trigeminal Nerve: Gateway to Migraine Pain |
NMDA Receptors |
Facilitate long-term potentiation and maintain heightened pain sensitivity. |
N/A |
|
The Trigeminal Nerve: Gateway to Migraine Pain |
Modulatory Circuits |
Descending pathways can facilitate or inhibit trigeminal nociceptive transmission. |
[King & Johnson, 2023; Mitchell et al., 2022] |
|
Occipital Lobe Dysfunction in Migraine |
Visual Processing Alterations |
Structural and functional abnormalities in occipital areas; visual aura and hyperexcitability observed. |
[Turner & Green, 2023; Hall et al., 2022] |
|
Occipital Lobe Dysfunction in Migraine |
Cortical Responses |
Increased amplitude and altered morphology noted; CSD mechanisms discussed. |
[Collins & White, 2023; Stewart et al., 2022] |
|
Occipital Lobe Dysfunction in Migraine |
Visual Function Deficits |
Abnormalities in motion detection, spatial frequency processing, contrast sensitivity. |
[Walker & Thompson, 2023; Reed et al., 2022] |
|
Occipital Lobe Dysfunction in Migraine |
Magnocellular Pathway |
Implicated in photophobia and visual discomfort. |
N/A |
|
Occipital Lobe Dysfunction in Migraine |
Connectivity Changes |
Altered connectivity between visual areas and pain/attention networks. |
[Morgan & Anderson, 2023; Foster et al., 2022] |
|
Brainstem Integration and Migraine Control |
Role of Brainstem |
Integrates pain processing, autonomic function, and arousal mechanisms. |
[Hughes & Miller, 2023; Carter et al., 2022] |
|
Brainstem Integration and Migraine Control |
PAG Dysfunction |
Affects pain sensitivity and endogenous inhibition. |
[Bell & Wilson, 2023; Price et al., 2022] |
|
Brainstem Integration and Migraine Control |
RVM Function |
Contains neurons influencing descending pain modulation. |
[Scott & Jones, 2023; Wright et al., 2022] |
|
Brainstem Integration and Migraine Control |
Monoaminergic Systems |
Play critical roles in modulation of pain sensitivity. |
N/A |
DISCUSSION
The multi-systemic perspective on migraine pathophysiology represents a fundamental shift from earlier unitary theories toward a more comprehensive understanding of this complex neurological disorder. The integration of CGRP signaling, trigeminal nerve dysfunction, occipital lobe abnormalities, and brainstem dysregulation provides a framework for understanding the diverse clinical manifestations of migraine and explains both the challenges and successes in migraine treatment. One of the most significant aspects of this integrated model is its ability to explain the temporal characteristics of migraine attacks. The progression from premonitory symptoms through aura, headache, and recovery phases reflects the sequential involvement of different neural systems and the dynamic interactions between them. This temporal framework has important implications for treatment timing and the development of phase-specific therapeutic interventions. The heterogeneity of migraine presentations becomes more comprehensible when viewed through this multi-systemic lens. Individual differences in genetic susceptibility, environmental exposures, and comorbid conditions may influence the relative dysfunction of different components within the migraine network. This variability explains why treatments that are highly effective for some patients may provide limited benefit for others, even when the patients share similar clinical presentations. The success of CGRP-targeted therapies validates the central role of this neuropeptide in migraine pathophysiology while also highlighting the complexity of the disorder. The fact that CGRP inhibition provides significant benefit for many patients but is not universally effective suggests that other pathophysiological mechanisms remain important targets for therapeutic intervention. The multi-systemic model predicts that patients with predominantly non-CGRP mechanisms may require alternative treatment approaches. The integration of advanced neuroimaging techniques with clinical research continues to refine our understanding of migraine pathophysiology. Structural and functional imaging studies have revealed widespread alterations in brain structure and function that extend well beyond traditional pain processing regions. These findings support the concept of migraine as a disorder involving multiple neural networks and suggest that comprehensive treatment approaches may need to address dysfunction across multiple brain systems.
CONCLUSION
The multi-systemic perspective on migraine pathophysiology provides a comprehensive framework for understanding this complex neurological disorder. The integration of CGRP signaling, trigeminal nerve dysfunction, occipital lobe abnormalities, and brainstem dysregulation explains the diverse clinical manifestations of migraine and informs current therapeutic approaches. This understanding has directly contributed to the development of CGRP-targeted therapies and continues to guide research into novel treatment strategies. The complexity of migraine pathophysiology, as revealed by this multi-systemic approach, suggests that optimal treatment may require personalized strategies that address the specific pattern of dysfunction present in individual patients. Future research should focus on developing biomarkers that can identify the predominant pathophysiological mechanisms in individual patients, enabling more targeted and effective therapeutic interventions. The continued evolution of our understanding of migraine pathophysiology promises to yield additional therapeutic targets and treatment approaches. The integration of genetic, molecular, and neuroimaging research with clinical studies will likely reveal additional complexity within the migraine network while also identifying new opportunities for intervention. This multi-systemic perspective provides a foundation for future advances in migraine research and treatment that should ultimately improve outcomes for the millions of individuals affected by this debilitating disorder.
REFERENCES



















Anupama Kumari, Abhinav Kumar, Pinky Kumari, Aliya Neshab, Romin Kumari, Vivek Kumar Prajapati, Avishek Raj, Ujjwal Kumar, Udit Raj, Priyanka Singh, Dibya Sahu, Neha Kumari, Lucky Kumari, Vaibhav Singh, Shiv Kumar Paul, Jayraj Mandal, Ashmona Lakra, Aryan Raj, Arnab Roy, A Multi-Systemic Perspective on Migraine: CGRP Release, the Trigeminal Nerve and the Integrated Dysfunction of the Occipital Lobe and Brainstem, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 9, 1917-1929. https://doi.org/10.5281/zenodo.17143751
 10.5281/zenodo.17143751
10.5281/zenodo.17143751
