1Head of Department of Pharmacy Practice, Channabasweshwar Pharmacy College (Degree), Latur, Maharashtra, India 413512.
2Research Scholar, Department of Pharmacology, Channabasweshwar Pharmacy College (Degree), Latur, Maharashtra, India 413512.
3,4Research Scholar, Department of Pharmacology, Channabasweshwar Pharmacy College (Degree), Latur, Maharashtra, India 413512.
Type 2 diabetes mellitus (T2DM) is a complex metabolic disease strongly influenced by alterations in the gut microbiota. Recent evidence shows that gut dysbiosis characterized by reduced microbial diversity, loss of short-chain fatty acid-producing bacteria, and enrichment of pro-inflammatory taxa-plays a significant role in insulin resistance, chronic inflammation, and impaired glucose metabolism. Key microbial signatures in T2DM include reduced Akkermansia, Faecalibacterium, Roseburia, and Bacteroides, along with increased Proteobacteria and opportunistic pathogens such as Escherichia–Shigella. These compositional shifts contribute to metabolic endotoxemia, oxidative stress, and disrupted gut barrier integrity. Metabolites such as SCFAs, bile acids, branched-chain amino acids, and lipopolysaccharides (LPS) influence multiple host pathways regulating glucose homeostasis, lipid metabolism and immune responses. Diet, probiotics, prebiotics, polyphenols, and herbal formulations can modulate gut microbiota composition and improve metabolic outcomes. Dysbiosis is also implicated in major microvascular complications, including diabetic nephropathy, neuropathy, and retinopathy, as well as macrovascular disease. Understanding the gut–host metabolic axis offers new opportunities for early diagnosis, prevention, and therapeutic intervention in T2DM. This review summarizes current evidence on microbiota alterations in T2DM, underlying mechanisms, links with complications, and microbiota-directed therapeutic strategies.
Diabetes mellitus (DM) is a chronic metabolic disease marked by persistently high blood sugar levels, resulting from impaired insulin secretion, reduced insulin sensitivity, or both, which disrupt normal glucose uptake and metabolism in tissues. Dysbiosis of the gut microbiota can contribute to insulin resistance and chronic low-grade inflammation, affecting glucose homeostasis and promoting the development and progression of type 2 diabetes mellitus (T2DM) (Yu Y et al., 2025). The gut microbiota, a complex microbial community in the gastrointestinal tract, plays a key role in metabolic health. Dysbiosis characterized by a higher Firmicutes/Bacteroidetes ratio, more Proteobacteria, reduced Akkermansia, and increased Prevotella and Bacteroides is associated with insulin resistance and T2DM. These microbial changes influence through its involvement in nutrient absorption, energy balance, and immune modulation (Li X et al., 2017). According to the International Diabetes Federation (IDF, 2025), approximately 589 million adults globally, aged 20–79 years, are living with diabetes. This number is projected to exceed 640 million by 2030 and 783 million by 2045 with the most rapid growth occurring in low- and middle-income countries (IDF, 2025). The gut microbiota influences the development and progression of T2DM through multiple mechanisms. Fermentation of dietary fibers by gut microbes produces short-chain fatty acids (SCFAs), which maintain gut barrier integrity and regulate host metabolism (Mashal R et al., 2024). Dysbiosis can cause metabolic endotoxemia and chronic inflammation, impairing insulin signaling. Additionally, gut microbes modulate bile acid signaling and SCFA-mediated cellular metabolism, affecting glucose homeostasis and insulin sensitivity (Fu Y et al., 2023). Diet and lifestyle significantly influence gut microbiota composition and function, while interventions such as prebiotics, probiotics, and polyphenol can restore microbial balance and improve glucose metabolism (Li Guang et al.,2023). Herbal medicines such as Gegen Qinlian Decoction, polysaccharides, and polyphenols are known to modulate gut microbiota by promoting beneficial bacteria and suppressing harmful microbes, which may contribute to their therapeutic effects (Gao Z et al., 2024). Growing evidence shows that gut microbiota dysbiosis contributes significantly to type 2 diabetes and its complications. Poorly controlled diabetes leads to metabolic disturbances such as dyslipidemia, oxidative stress, and hypertension that drive microvascular complications such as nephropathy, neuropathy, retinopathy and macrovascular complications like cardiovascular disease, Overall, the link between gut microbiota imbalance, metabolic dysfunction, and inflammation highlights microbiota modulation as a promising strategy for managing diabetes and its complications (Camelia O Iatcu et al., 2022). 16S rRNA gene sequencing is a molecular technique used to identify and classify bacteria in complex ecosystems such as the human gut. By targeting conserved and variable regions of the 16S rRNA gene, it allows researchers to determine microbial composition and abundance from fecal or tissue samples. In type 2 diabetes, this method has revealed gut dysbiosis, including reduced diversity and lower levels of beneficial short-chain fatty acid-producing bacteria like Roseburia, Faecalibacterium, and Akkermansia. These microbial changes are linked to metabolic dysfunction, chronic inflammation, and impaired gut barrier integrity, contributing to the onset and progression of diabetes and its complications (Wang Y et al., 2023). Diabetic individuals show reduced microbial diversity, altered gut bacterial ratios, and disrupted short-chain fatty acid production, affecting inflammation, insulin sensitivity, and glucose regulation.
2. Composition of gut microbiota in T2DM
In 2001, Joshua Lederberg, a former Nobel Prize winner, first defined the ‘human microbiome’ as an ecological community of commensal, symbiotic and pathogenic microorganisms. The first study on gut microbiota of T2DM was reported in 2010 (N. Larsen et al., 2010). There are two most common forms of diabetes, namely, type 1 diabetes mellitus and type 2 diabetes mellitus and it account for about 90% of all diabetes cases, resulting from insulin resistance combined with impaired insulin secretion. The gut microbiota of individuals with T2DM shows characteristic alterations at both phylum and genus levels (Qin J. et al., 2012). A marked decline in Bacteroidetes, along with an increase in Proteobacteria and Actinobacteria, is commonly observed. At the genus level, beneficial SCFA-producing bacteria such as Bacteroides, Blautia, Faecalibacterium, Lachnospira, Pseudobutyrivibrio, and Roseburia are significantly reduced, while opportunistic or mucin-degrading genera including Escherichia-Shigella, Subdoligranulum, Akkermansia, Enterococcus, Bifidobacterium, Klebsiella and Lactobacillus tend to be enriched in T2DM (Ejtahed HS et al., 2020). Bacteroides has strong genomic potential for fermenting diverse carbohydrates into short chain fatty acid (SCFAs), a function essential for metabolic balance, yet its reduction in diabetes contributes to impaired glucose homeostasis. Escherichia-Shigella is elevated in T2DM and acts as an opportunistic pathogen, releasing pro-inflammatory molecules like lipopolysaccharide (LPS) and peptidoglycan (PGN) that disrupt epithelial integrity and promote low-grade inflammation (Douglas J. Morrison et al., 2016). Lachnospiraceae was a kind of anaerobic, spore-forming bacteria, which helping the host to digest some carbohydrates and fibers. Akkermansia muciniphila sensitive indicator of intestinal permeability, improving glucose tolerance. obesity and dyslipidemia were related to the low prevalence of phylum Bacteroidetes and the increase of Firmicutes/Bacteroidetes ratio. The ratio between Firmicutes/Bacteroidetes is often used as an indicator of gut microbial balance and has been associated with metabolic health (Xue Zhao et al., 2020). Gut microbiota consists of more than 1000 bacterial species, mainly distributing in 9 phyla, most of which belong to the Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria. Betaproteobacteria was highly increased and positively associated with plasma glucose. Alterations in gut microbial ratios particularly higher Bacteroidetes/Firmicutes ratios, show a positive correlation with plasma glucose, indicating a clear association between intestinal microbiota composition and T2DM. These microbial shifts influence T2DM progression through several metabolic pathways, particularly those involving SCFAs, bile acids, BCAAs, imidazole propionate, and LPS (Lili Zhang et al., 2021).In diabetic individuals, key butyrate-producing bacteria including Faecalibacterium prausnitzii, Roseburia intestinalis, Eubacterium rectale, and related Clostridiales species are consistently reduced. Conversely, opportunistic pathogens such as Bacteroides caccae, Clostridium hathewayi, Eggerthella lenta, Clostridium ramosum, and Escherichia coli, along with mucin-degrading Akkermansia muciniphila and sulfate-reducing Desulfovibrio species, are enriched. Lactobacillus shows a positive association with fasting blood glucose and glycated hemoglobin (HbA1c), whereas Clostridium exhibits negative correlations with glycemic and lipid markers (Eline Birkeland et al., 2020). This study investigated gut microbiome differences among Chinese hypertension patients with and without T2DM. Using 16S rRNA sequencing on fecal samples from 72 hypertension patients. Healthy controls showed higher microbial diversity, while hypertensive and HT-T2DM groups displayed distinct shifts in key taxa, including reduced Bacteroides and Blautia and increased Prevotella copri and Klebsiella pneumoniae (Hongying Ding et al., 2023). A cross-sectional study of a large, population-based sample found associations between gut microbiome composition and both type 2 diabetes prevalence and insulin resistance, and further showed that a higher abundance of 12 specific taxa Christensenellaceae, Clostridiaceae, Peptostreptococcaceae, Christensenellaceae R7 group, Marvinbryantia, Ruminococcaceae, Clostridium sensu stricto, Intestinibacter, and Romboutsia may be beneficial in reducing the risk of insulin resistance and T2DM. Butyrate has been suggested to exert beneficial metabolic effects by enhancing mitochondrial activity, improving overall energy metabolism, activating intestinal gluconeogenesis, and preventing metabolic endotoxemia and inflammation through multiple pathways involving gene expression and hormone regulation (Zhangling Chen et al., 2021). For mild obesity-related diabetes (MOD), Class Clostridia and Order Clostridiales appeared to have protective effects, suggesting that higher levels of these bacteria may help reduce MOD risk, whereas the bacterial species Catus was associated with an increased risk. In severe insulin-resistant diabetes (SIRD), the genus Holdemania was linked to a higher likelihood of developing the condition. Additional microbial taxa were also associated with severe insulin-deficient diabetes (SIDD) and mild age-related diabetes (MARD) (Yasser Morsy et al., 2025). The composition and activity of gut microbes, as well as the metabolites they produce such as short-chain fatty acids, lipopolysaccharides, trimethylamine N-oxide, and bile acids can significantly impact diabetes development. A healthy gut microbiota (eubiosis) produces essential short-chain fatty acids such as butyrate, acetate, and propionate through the fermentation of dietary fiber, which act on GPR41 and GPR43 receptors to maintain colonocyte metabolism, strengthen the gut barrier, reduce inflammation and support insulin secretion. This balanced microbial environment also stimulates the release of GLP-1 and PYY, improving appetite (Yifeng Fu et al., 2023). Firmicutes make up the greatest proportion of the gut microbiota (64%), followed by the Bacteroidetes (23%), Proteobacteria (8%) and lastly Actinobacteria (3%) (Hattori M et al., 2009).
2.1 Alteration of gut microbiota in T2DM
|
Sr. no. |
Gut microbiome |
Direction in T2DM |
Role / Findings in T2DM |
References |
|
|
1 |
Akkermansia muciniphila |
|
Reduces levels associated with impaired mucin layer, increased inflammation, insulin resistance. |
(Li J et al., 2023) |
|
|
2 |
Faecalibacterium prausnitzii |
|
Loss of this major butyrate producer contributes to gut inflammation and metabolic dysregulation. |
(Sikalidis AK et al., 2020) |
|
|
3 |
Roseburia spp. (butyrate producers) |
↓ Decreased |
Low butyrate production contributes to reduced GLP-1 release and insulin resistance. |
(Mohammed A S et al., 2025) |
|
|
4 |
Bifidobacterium spp. |
↓ Decreased |
Reduced levels linked to higher LPS load, impaired gut barrier, and chronic inflammation in T2DM. |
(Ecklu-Mensah G et al., 2023) |
|
|
5 |
|
↑ Increased |
Enrichment of endotoxin-producing strains contributes to systemic inflammation → insulin resistance. |
(Li J et al., 2023) |
|
|
6 |
Firmicutes/Bacteroidetes ratio |
Altered |
T2DM associated with disrupted ratio, reduced SCFAs, and altered energy harvesting. |
(Mohammed A S et al., 2025) |
3. Mechanism linking gut microbes to T2DM
Dietary fiber fermentation:
Gut microbes play a key role in type 2 diabetes by regulating glucose metabolism, maintaining gut barrier integrity, modulating inflammation, influencing bile acid and hormonal signaling and producing metabolites that affect insulin sensitivity. Microbial metabolites produced during anaerobic fermentation in the gut, including short-chain fatty acids (SCFAs), branched-chain amino acids (BCAAs), succinate, indole, and imidazole, act as key mediators in microbe-to-host communication (Stacey M Lambeth et al., 2015). These metabolites are primarily generated by gut microbial genera including Akkermansia, Prevotella, Ruminococcus, Coprococcus, Faecalibacterium, Eubacterium, Roseburia, Clostridium, Bacteroides, Lactobacillus, Streptococcus, Propionibacterium, and Fusobacterium (Hideo Ohira et al., 2017). Butyrate producing microbes are notably depleted in patients with T2DM, the Clostridiales order, genera Ruminococcus and Subdoligranulum, and species including Eubacterium rectale, Faecalibacterium prausnitzii, Roseburia intestinalis, and Roseburia inulinivorans. Additionally, the genera Bacteroides, Prevotella, and Bifidobacterium are also present in significantly lower abundance in individuals with T2DM. Bifidobacterium species are recognized for their beneficial effects on health, particularly their capacity to strengthen intestinal barrier integrity, which helps lower circulating endotoxin levels and systemic inflammation. The genus Akkermansia plays a critical role in maintaining the integrity of the mucin layer and reducing inflammation and Faecali prausnitzii is a secondary reduction in systemic inflammation and an improvement in insulin resistance (A. L. Cunningham et al., 2021). Dysbiosis alters the production of key microbial metabolites such as short-chain fatty acids (SCFAs). The lipopolysaccharides (LPS) and aromatic amino acids, which collectively promote chronic low-grade inflammation, increased intestinal permeability, and insulin resistance. These disturbances contribute not only to the onset of T2DM but also to the progression of its microvascular complications, including diabetic retinopathy, nephropathy, and neuropathy, through mechanisms involving oxidative stress, vascular dysfunction, and immune dysregulation (Daniela Maria Tanase et al., 2020).
Short chain fatty acid and gut barrier maintenance:
Higher amounts of Betaproteobacteria and Lactobacillus have also been associated with increased blood glucose levels. Higher abundances of Lactobacillus gasseri and Streptococcus mutans along with reductions in Roseburia intestinalis and Faecalibacterium prausnitzii have been observed in T2DM. Studies report that, people with metabolic problems often have higher levels of Lactobacillus species and increased microbial oxidative-stress activity which have been linked to insulin resistance, B-cell dysfunction and impaired glucose tolerance. The genera Clostridium and Phascolarctobacterium are generally linked to better insulin sensitivity, while Dialister is linked to poorer insulin sensitivity (E. Wright Jr et al., 2006). Short-chain fatty acids (SCFAs) such as acetate and propionate act as metabolic substrates that support lipogenesis and gluconeogenesis in the liver and other peripheral tissues. In contrast, butyrate primarily serves as a major energy source for colonic epithelial cells and has been reported to stimulate intestinal gluconeogenesis in both the small intestine and colon. Short chain fatty acid also regulates the release of several gut-derived peptide hormones, including GLP-1, gastric inhibitory peptide (GIP), GLP-2, and PYY, which collectively contribute to improved insulin sensitivity, reduced inflammatory responses, and enhanced satiety (Gwen Tolhurst et al., 2012).
Metabolic endotoxemia and inflammation: Chronic low-grade inflammation observed in obesity and type 2 diabetes is largely driven by elevated levels of pro-inflammatory cytokines such as TNF-α and the interleukins IL-1 and IL-6. Disruptions in gut microbial composition, known as dysbiosis, can promote closer bacterial contact with the intestinal epithelium, thereby initiating inflammatory signaling. Dietary supplementation with selected probiotic strains such as Lactobacillus, Bifidobacterium, Clostridium, and Akkermansia has been reported to reduce inflammatory pathways and enhance insulin sensitivity (Torsten P. M. Scheithauer et al., 2020). Increased gut permeability is a key driver of inflammation in obesity and type 2 diabetes and is often linked to high-fat diet intake. When the intestinal mucus layer is disrupted, bacterial components such as lipopolysaccharide (LPS) and flagellin can reach the epithelium and activate inflammatory pathways. Reductions in beneficial species such as Akkermansia muciniphila are associated with impaired mucosal integrity and worsened insulin resistance. Probiotic and prebiotic interventions can restore barrier function and reduce gut permeability, highlighting the essential role of the gut microbiota in modulating inflammation and type 2 diabetes mellitus (Paola Brun et al., 2006).
Bile acid signaling:
Bile acids regulate glucose metabolism, with gut microbes converting primary bile acids such as cholic acid, chenodeoxycholic acid into secondary bile acids like deoxycholic acid, lithocholic acid. Deconjugation by microbes such as Lactobacillus, Bifidobacterium, Bacteroides, and Clostridium allows further colonic metabolism. Primary bile acids act via farnesoid X receptor (FXR), while secondary bile acids signal through Takeda G-protein-coupled receptor (TGR5), promoting GLP-1 secretion and improving glycemic control. Cholic acid reduces triglycerides and prevents insulin resistance in high-fat diet models. Clinically, bile acid sequestrants like colesevelam improve glucose and lipid profiles, likely through GLP-1. Benefits of Roux-en-Y gastric bypass may also involve Akkermansia muciniphila–mediated bile acid changes, whereas pollutants can disrupt bile acid metabolism and impair glucose homeostasis (Annemarie B et al., 2015).
Short-chain fatty acids and cellular metabolism:
Lipopolysaccharide (LPS), which comprises lipids and polysaccharides, is a major component of the cell wall of gram-negative bacteria. Lipopolysaccharide (LPS), a key component of Gram-negative bacterial cell walls, triggers innate immune responses by activating TLRs on macrophages and neutrophils, leading to the release of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 via NF-κB and MAPK pathways (Liu T et al., 2012). Short-chain fatty acids (SCFAs) like acetate, propionate, and butyrate can attenuate LPS-induced inflammation through activation of GPCRs (FFAR2, FFAR3, GPR109A) and inhibition of histone deacetylases (HDACs), reducing pro-inflammatory cytokine production and promoting anti-inflammatory mediators such as IL-10. Thus, SCFAs play a protective role against LPS-mediated inflammatory responses (J. Czepiel et al., 2014). Short-chain fatty acids (SCFAs) regulate glucose metabolism through multiple mechanisms. They activate G-protein-coupled receptors such as FFAR2 (GPR43) and FFAR3 (GPR41), stimulating the secretion of intestinal hormones GLP-1 and PYY, which enhance insulin secretion, reduce glucagon release, improve glucose uptake in muscle and adipose tissue, and promote satiety (SCFAs also increase the expression and membrane translocation of GLUT4 in skeletal muscle, facilitating glucose absorption (Portincase p et al., 2023). Additionally, SCFAs activate AMP-activated protein kinase (AMPK) by increasing the cellular AMP/ATP ratio, leading to enhanced glucose transport, increased fatty acid oxidation, and inhibition of gluconeogenesis through down regulation of G6Pase and PEPCK in the liver. Together, these pathways allow SCFAs to improve blood glucose homeostasis and energy metabolism (Jin He et al., 2020). Short-chain fatty acids (SCFAs) regulate lipid metabolism through multiple mechanisms. They are converted into acetyl-CoA, serving as a substrate for fatty acid and triglyceride synthesis or entering the TCA cycle for energy production. SCFAs activate AMP-activated protein kinase (AMPK), which enhances PGC-1α (Peroxisome Proliferator-Activated Receptor Gamma Co-activator 1-alpha) expression and activates transcription factors PPARα/γ, promoting fatty acid oxidation and reducing triglyceride and free fatty acid levels. AMPK activation also regulates HSL (Hormone-Sensitive Lipase) and ATGL (Adipose Triglyceride Lipase) to modulate lipolysis. Additionally, SCFAs upregulate uncoupling proteins (UCP1, UCP2, UCP3), increasing thermogenesis and mitochondrial fatty acid oxidation, while enhancing adiponectin signaling to improve lipid utilization. Collectively, these pathways allow SCFAs to reduce lipid accumulation, increase energy expenditure, and maintain metabolic homeostasis (Carmen, G et al., 2006).
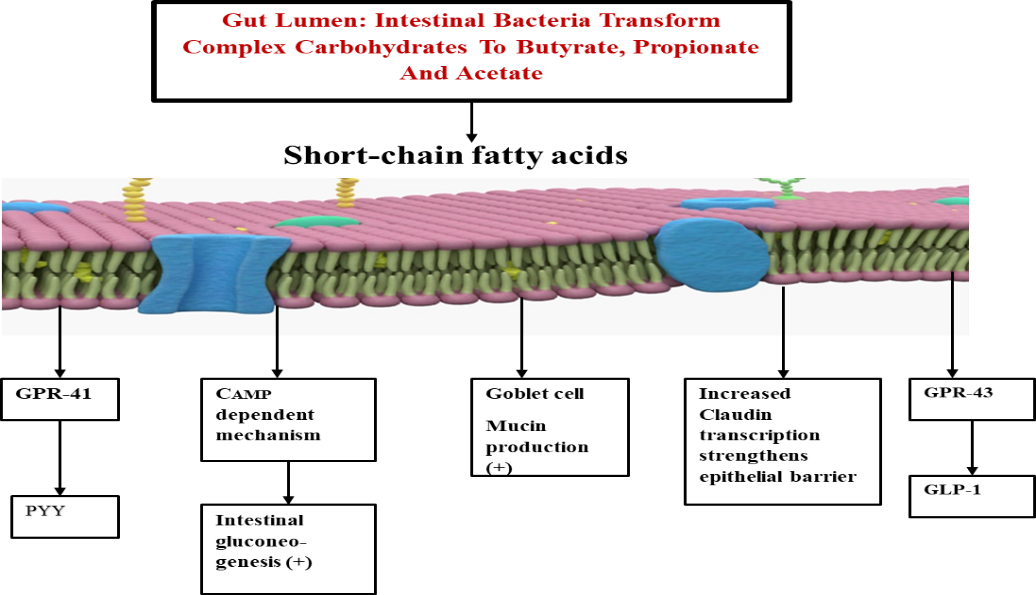
Figure 1. The effects of short chain fatty acids (SCFAs): Peptide YY (PYY) is expressed by short-chain fatty acid (SCFA)–dependent activation of the G-protein-coupled receptor GPR41, while glucagon-like peptide-1 (GLP-1) is expressed after SCFA-dependent activation of GPR43. Both GLP-1 and PYY inhibit gut motility, suppress appetite, and decrease energy harvest. Propionate and butyrate stimulate intestinal gluconeogenesis through CAMP dependent mechanisms. Additionally, butyrate strengthens the endothelial and intestinal barriers by reducing molecular permeability and up regulating Claudin-1 transcription.
4. Diet, lifestyle and microbiota modulation
Diet has a major impact on the gut microbiota, and even short-term changes can alter these microbial communities within 24 hours, though the effects are reversible. Gut microbes help the body by producing vitamins, essential amino acids, and important metabolites. They also break down undigested food to form short-chain fatty acids (SCFAs) such as butyrate, propionate, and acetate, which provide energy to intestinal cells and help maintain metabolic health. Different dietary components proteins, fats, carbohydrates, probiotics, and polyphenols can influence the types of microbes present and affect immune and metabolic functions (Siobhan F Clarke et al., 2016). Dietary fats and carbohydrates strongly shape the gut microbiota. Saturated fats tend to increase total anaerobic bacteria, especially Bacteroides and Bilophila, while unsaturated fats cause broader changes in the overall microbial community. Both digestible and non-digestible carbohydrates increase Bifidobacterium and reduce Clostridia, and non-digestible carbohydrates additionally promote beneficial groups like Lactobacillus, Ruminococcus, Eubacterium rectale, and Roseburia. Probiotics and polyphenols also support the growth of Bifidobacterium and other lactic acid producing bacteria while lowering harmful Clostridia species (Rasnik K. Singh et al., 2017). Clinical evidence suggests that dietary fibers, digestible and non-digestible carbohydrates including arabinoxylans, galacto-oligosaccharides, inulin, and oligofructose, promote a range of beneficial bacteria and enhance the growth of beneficial gut bacteria while suppressing potentially harmful taxa. Similarly, polyphenols modulate gut microbial composition by inhibiting pathogenic organisms such as Helicobacter pylori and Staphylococcus spp. and by promoting the proliferation of beneficial genera like Lactobacillus and Bifidobacterium (Shanthi G. Parkar et al., 2008). Flavonoids, phenolic acids, and other polyphenols such as anthocyanins, quercetin, rutin, chlorogenic acid, and caffeic acid can increase beneficial gut bacteria like Bifidobacterium and Lactobacillus while reducing harmful species. Catechins also promote the growth of the Clostridium coccoides Eubacterium rectale group, Bifidobacterium, and E. coli, and suppress the Clostridium histolyticum group, showing their selective effects on gut microbiota (Maria V. Salma et al., 2009). Polyphenols are fermented by the gut microbiota to increase beneficial bacteria such as Bifidobacteria and Lactobacilli, lower the Firmicutes/Bacteroidetes ratio, and enhance short-chain fatty acid production. They also promote the growth of Akkermansia and key butyrate-producing species like Faecalibacterium prausnitzii and Roseburia, highlighting their positive role in gut microbial balance and metabolic health (Qi Yang et al., 2020). Gut microbiota are capable of synthesizing several vitamins, including vitamin K and B-group vitamins. They can influence the gut microbial communities. Vitamin A deficiency has been associated with reduced microbial diversity, characterized by a decline in butyrate-producing bacteria such as Escherichia coli and Clostridium butyricum, along with an increase in opportunistic pathogens like Enterococcus. Vitamins C, D, and E also promote health-associated microbes, especially Bifidobacterium and Lactobacillus. Supplementation with retinoic acid has been shown to enhance the abundance of beneficial genera, including Allobaculum, Aggregatibacter, Bifidobacterium, Dialister, and Enhydrobacter (Renee Wilson et al., 2018).
5. Impact of Herbal Medicines on Gut Microbiota
Herbal medicine uses medicinal plants and their active compounds to help manage diseases. In type 2 diabetes (T2DM), herbal treatments may improve gut microbiota balance and support better glucose control. Plant polyphenols, which have strong antioxidant and anti-inflammatory properties, can positively influence key gut microbes. These effects help improve insulin sensitivity, stabilize blood sugar levels, and lower blood lipids, making polyphenols beneficial for T2DM management (Andy W.C. Man et al., 2020). Polyphenols help shape the gut microbiota by increasing beneficial bacteria like Bifidobacterium, Akkermansia, and Lactobacillus, while reducing harmful Proteobacteria. These changes support gut balance and improve metabolic health. Polyphenols also act directly on organs that control glucose and lipid metabolism, helping improve obesity and type 2 diabetes. Additionally, short-chain fatty acids like butyrate strengthen the gut barrier and reduce inflammation by promoting M2-like macrophages and lowering cytokines such as TNF-α, IL-1β, and IL-6 (Keyu Chen et al., 2022). Chinese herbal monomers contain key bioactive constituents such as polysaccharides, flavonoids, alkaloids, and saponins, which collectively contribute to the regulation of gut microbiota and improvement of metabolic function in diabetes. These compounds enhance insulin sensitivity and glucose metabolism by modulating microbial composition and promoting beneficial metabolites. Notably, polysaccharides from Cyclocarya paliurus and Astragalus membranaceus have been shown to increase SCFA-producing bacteria, elevate SCFA levels, and stimulate the expression of GLP-1 and PYY, thereby improving glucose tolerance and supporting glycemic control in type 2 diabetes (Jing Liu et al., 2025). Gegen Qinlian Decoction (GQD) is a well-studied anti-hyperglycemic herbal formulation with demonstrated regulatory effects on the gut microbiota. Animal studies show that GQD enhances bile acid metabolism and activates the TGR5/cAMP/PKA/CREB signaling pathway, leading to increased GLP-1 secretion. These actions collectively reshape gut microbial structure, reduce inflammatory cytokines, and lower blood glucose levels. Furthermore, experimental evidence suggests that GQD ameliorates hyperglycemia and protects pancreatic function by modulating gut microbiota and their metabolic products (Zheng Yujiao et al., 2020). Qinglian Hongqu Decoction and Jiang Tang Sanhuang (JTSH) Pill have been shown to modulate metabolic pathways by activating the intestinal FXR/FGF15 and TGR5/GLP-1 signaling axes, resulting in reduced lipid accumulation and improved insulin sensitivity. Clinical evidence supports these findings, as a retrospective cohort study reported that one year of JTSH tablet treatment lowered the risk of poor glycemic control by 17%. Patients with T2DM also demonstrated significant reductions in blood glucose and insulin resistance, alongside enhanced pancreatic β-cell function, indicating the therapeutic potential of JTSH in metabolic regulation (Chester Yan Jie Ng et al., 2022).
6. Gut microbiota with diabetes complications
6.1 Diabetic nephropathy: Diabetic nephropathy (DN), a major microvascular complication affecting nearly 40% of individuals with diabetes, substantially increases the risk of renal failure and contributes to the global disease burden. Recent evidence indicates that gut microbiota and their metabolites influence DN progression via the gut–kidney axis. Dysbiosis can compromise intestinal barrier integrity, allowing endotoxins and pathogens to translocate into the circulation, thereby triggering inflammation and oxidative stress that accelerate renal injury (Salman Hussain et al., 2019). Recent studies show that the gut microbiota plays an important role in type 2 diabetes mellitus and in complications such as diabetic nephropathy. Research also suggests that the gut and kidneys can affect each other through changes in metabolism, immunity and hormones, a connection known as the “intestinal–renal axis.” In patients with diabetic nephropathy, several studies have reported specific changes in gut microbes, including higher levels of Clostridium and Aspergillus and lower levels of Rhodococcus (Maria v. Salguero et al., 2019). Studies also show that several factors can damage the intestinal lining in individuals with diabetic nephropathy, making it more permeable. This increased permeability allows metabolites such as indole and p-cresol to pass into the bloodstream, where they can contribute to kidney injury. Patients with diabetic nephropathy often exhibit systemic inflammation and weakened innate immune responses, which may further worsen the disease (Tian En et al., 2023). Research shows that the gut microbiota modulates antigen responsiveness within lymphoid tissues and plays a central role in the initiation and maturation of intestinal immunity. Key genera including Bacteroides, Bifidobacterium, Lactobacillus, and Bacillus proteus are particularly important for supporting immune function. Conversely, disturbances in microbial composition can impair macrophage maturation and enhance TNF-α and IL-6 release following TLR activation, promoting renal inflammation. Such dysbiosis may also heighten TLR-2 and TLR-4 signalling and elevate pro-inflammatory cytokine production, contributing to immune dysregulation and renal damage in diabetic nephropathy (Jiang Haiyan et al., 2023). An increased abundance of urease-producing bacteria raises intestinal pH, which enhances the translocation of uremic toxin precursors such as cresol, indole, and trimethylamine into the bloodstream, thereby promoting uremic toxin formation. These toxins subsequently induce oxidative stress and drive tubulointerstitial fibrosis, contributing to the progressive deterioration of renal function (Yongxiu Jin et al., 2023). Diabetic nephropathy is marked by glomerular basement membrane thickening, mesangial expansion, podocyte loss, glomerulosclerosis, and tubulointerstitial fibrosis, culminating in progressive albuminuria and reduced glomerular filtration. Chronic hyperglycemia promotes the accumulation of advanced glycation end products (AGEs) in endothelial cells, the glomerular basement membrane, and podocytes, which directly disrupt glomerular structure and function and activate growth factors and cytokines via RAGE, thereby contributing to proteinuria (Chenhua Wu et al., 2022).The increase of intracellular blood glucose level leads to the increase of diacylglycerol (DG) synthesis and activation of PKC, thus inducing the production of the signal molecules such as endothelin-1 (ET-1), transforming growth factor-β1 (TGF-β1), interleukin-1 (IL-1) and reactive oxygen species (ROS) in kidney cells. Consequently, the increase in ET-1 results in the injury of endothelial cells. The overproduction of TGF-β1 leads to the accumulation of glomerular extracellular matrix and the hypertrophy of kidney cells (Kanwar Yashpal S et al., 2011). Additionally, the glucose can be converted into sorbitol by aldose reductase (AR) via polyol pathway under the diabetic circumstance. Sorbitol is prone to accumulate in kidney cells causing permeability alteration, cell swelling and rupture, which ultimately aggravate. The RAAS is over-activated, the ANG-II and aldosterone are increased under the hyperglycemic circumstance, which leads to the elevation of blood pressure and the destruction of sodium balance. Chronic glomerular hypertension, hyperperfusion and hyperfiltration result in glomerular sclerosis and renal interstitial fibrosis. In DN, increase ROS production due to hyperglycemia can cause oxidative stress, which can directly damage podocytes, mesangial cells and endothelial cells, and lead to albuminuria and tubulo-interstitial fibrosis. the alterations of gut microbial composition in DN are characterized by an increased abundance of pernicious bacteria such as Desulfovibrio and Coprobacillus, and the decrease in probiotics such as Lachnospira and Intestinibacter (Hui Zhao et al., 2023). Diabetic nephropathy is a common microvascular complication in patients with diabetes. the genera Lactobacillus, Clostridium IV, Paraprevotella, Clostridium sensu stricto, Desulfovibrio, and Alloprevotella were enriched in the fecal samples of patients with CKD, while Akkermansia and Parasutterella were enriched in those of NC subjects, (Xiao Lu et al., 2023).
6.2 Diabetic Neuropathy
Recent evidences suggest that association between diabetic peripheral neuropathy (DPN) and dysregulation of the gut microbiota. Several studies have demonstrated significant reductions in both bacterial α-diversity and β-diversity, indicating a loss of microbial richness and structural complexity in individuals with DPN. These alterations are accompanied by marked shifts in overall microbial community composition. Such gut microbial imbalance is thought to influence host neuroimmune pathways, particularly through the dysregulated release of pro-inflammatory mediators. These inflammatory signals can activate and sensitize nociceptive pathways, thereby contributing to peripheral nerve dysfunction and modulating pain perception in DPN (Jiayu Wu et al., 2020). Most of the relevant gut microbiota belong to the Firmicutes and Bacteroidetes phyla. Patients with diabetic neuropathy show an increased abundance of Firmicutes and a decreased abundance of Bacteroidetes, resulting in a higher Firmicutes/Bacteroidetes (F/B) ratio. This pattern is consistent with previous studies and has been linked to obesity-related metabolic changes. Elevated F/B ratios have also been implicated in the pathogenesis of neurological disorders, including neuropathic pain and cognitive impairment, suggesting a mechanistic role for gut dysbiosis in DPN (Amelie Joly et al., 2020).
Gut microbiota is increasingly recognized as a key regulator of cognitive function through the gut–brain axis, and its dysbiosis is known to promote neuroinflammatory processes. In DPN rat models, gut microbial profiles revealed a significant increase in the relative abundance of Escherichia–Shigella, Ruminococcaceae-UCG-010, Bifidobacterium, Parasutterella, Enterococcus, Anaeroplasma, Allobaculum, Turicibacter, Coprococcus-3, and Dubosiella. Conversely, the relative abundance of Ruminiclostridium-6, Parabacteroides, Lachnospiraceae, Anaerotruncus, and Rikenella was markedly reduced in DPN. Notably, Anaeroplasma, Turicibacter, and Coprococcus were significantly more abundant in the Control diabetic group compared to the non-control diabetic group, further indicating diet-related modulation of microbial signatures in DPN (Wei Huang et al., 2023). Imbalance of the intestinal microbiota can promote excessive production of amyloid, lipopolysaccharide (LPS), and other inflammation-related molecules, while simultaneously disrupting the intestinal mucosal barrier. These alterations facilitate microglial activation and trigger neuroinflammatory responses within the central nervous system, potentially contributing to neurodegeneration. The inflammatory cascade therefore represents a plausible mechanistic link between gut microbiota dysbiosis and CNS dysfunction. Overall, our findings indicate that persistent elevations in the abundance of Mycobacterium, Turicibacter, and Actinobacteria may play a contributory role in the development of DPN accompanied by cognitive impairment (Cattaneo A. et al., 2017).
6.3 Diabetic retinopathy
Diabetic retinopathy (DR), a major microvascular complication of diabetes and a leading cause of adult blindness, is increasingly linked to gut microbiota dysbiosis. Evidence within the “gut–retinal axis” framework shows that patients with DR exhibit distinct alterations in gut microbial composition and diversity. Such dysbiosis can impair intestinal barrier integrity, allowing microbial metabolites to enter systemic circulation and promote inflammation, thereby contributing to the onset and progression of DR (Thakur PS et al., 2022). Diabetic retinopathy (DR) is among the most prevalent microvascular complications of diabetes and remains a major global public health concern due to its high incidence and substantial risk of vision loss. As the leading cause of visual impairment and the foremost cause of blindness in adults, DR arises from a multifactorial pathogenic process. Its development reflects the interplay of microvascular dysfunction, retinal neurodegeneration, metabolic disturbances, genetic susceptibility, and dysregulated immune and inflammatory responses (Alarcon Yempen R et al., 2021). Gut microbiota dysbiosis can initiate both local and systemic inflammatory responses, thereby contributing to the development of diabetes and its microvascular complications, including the onset and progression of diabetic retinopathy (DR). Under diabetic conditions, alterations in gut microbial composition may provoke retinal neurovascular inflammation, driving structural and functional damage within the retina (Beli E et al., 2018).Diabetic retinopathy (DR) is increasingly recognized as a chronic metabolic–inflammatory disorder in which retinal neurons, vascular endothelial cells, and glial cells collectively constitute a highly integrated neurovascular unit. As DR advances, neurovascular coupling becomes progressively impaired and the integrity of the blood–retinal barrier (BRB) deteriorates. This disruption leads to the establishment of a persistent inflammatory microenvironment driven by complex interactions among activated immune cells, pro-inflammatory cytokines, and vascular components, ultimately resulting in microvascular dysfunction (Gardner TW et al., 2017). Microglia the resident innate immune cells of the retina play a central role in mediating both neurodegenerative and vascular injury in DR. They exhibit two major polarization states: the pro-inflammatory M1 phenotype and the anti-inflammatory, neuroprotective M2 phenotype. In the early stages of DR, microglia predominantly adopt the M2 phenotype, facilitating retinal protection through phagocytic clearance of debris and secretion of neurotrophic factors that support neuronal survival and tissue repair. As the disease progresses, however, microglia increasingly shift toward the M1 phenotype, characterized by heightened release of cytotoxic mediators such as interleukin-1 (IL-1) and tumour necrosis factor-α (TNF-α). This phenotypic transition amplifies inflammatory damage and contributes substantially to retinal neurovascular injury (Simo R, Stitt AW et al., 2018). At the phylum level, DR patients showed decreased Firmicutes and increased Bacteroidetes and Desulfobacterota compared with T2DM patients. Pasteurellaceae levels were reduced, while Eubacteriaceae showed an increasing trend. Genus-level analysis revealed a significant loss of anti-inflammatory bacteria (Faecalibacterium, Bifidobacterium, Ruminococcus, Turicibacter, Streptococcus, Lactobacillus, Butyricimonas) and an increased abundance of the pro-inflammatory genus Shigella, indicating a shift toward a pro-inflammatory gut profile in DR (Das T et al., 2021). Activation of Toll-like receptor 2 (TLR2) by Prevotella stimulates antigen-presenting cells to secrete IL-23 and IL-1, promoting Th17 cell-mediated inflammatory responses. The Th17 effector cytokine, interleukin-17A (IL-17A), induces retinal endothelial cell apoptosis via the IL-17A/IL-17 receptor (IL-17R) axis, involving sequential activation of the Act1 adaptor protein and Fas-associated death domain (FADD), which ultimately triggers caspase-dependent cell death. These findings suggest that Prevotella enrichment in DR patients may aggravate retinal injury through Th17-driven inflammatory mechanisms (Liqing Xie et al., 2019). At the genus level, DR patients exhibited enrichment of Faecalibacterium, Roseburia, Lachnospira, and Romboutsia, with a concurrent depletion of Akkermansia compared to healthy individuals. Relative to the DM group, DR patients showed increased abundance of five genera, including Prevotella, and decreased levels of Bacillus, Veillonella, and Pantoea. Animal studies indicate that intermittent fasting can prevent DR by reshaping the gut microbiota toward TUDCA-producing species, which confer retinal protection via activation of TGR5. These findings highlight TGR5 as a potential therapeutic target and suggest that gut microbial alterations may contribute to DR pathogenesis (Beli et al., 2019). DR patients had elevated Prevotella, Faecalibacterium, Subdoligranulum, Agathobacteria, and Olsenella and reduced Bacillus, Veillonella, and Pantoea abundances at the genus level Moreover, we found that Faecalibacterium and Lachnospira were depleted in DM patients compared with healthy controls at the genus level, and Klebsiella and Enterococcus were enriched (zhao et al., 2019). In people with diabetic retinopathy (DR), levels of Faecalibacterium and Roseburia in the gut are greatly reduced, similar to what has been seen in patients with ulcerative colitis. At the same time, DR patients tend to have higher levels of Prevotella and Subdoligranulum. Prevotella is known to be one of the main and common bacteria in the human gut. However, there is still no clear evidence showing that Subdoligranulum or Faecalibacterium directly contributes to the development of diabetic retinopathy (Zixi Zhou et al., 2021). Elevated lipopolysaccharide (LPS) levels in T2DM patients promote microbial translocation and activate the TLR4/MyD88–NF-κB pathway, leading to increased pro-inflammatory cytokines and impaired insulin signaling. This LPS-driven inflammation weakens the gut barrier, damages vascular endothelial cells, disrupts vascular function, and contributes to the worsening of diabetic retinopathy (DR) (Yining Zeng et al., 2025)
6.4 Cardiovascular complications
Cardiovascular disease (CVD) is a leading cause of illness and death around the world. with recognized risk factors including hypertension, dyslipidemia, smoking, and physical inactivity. Studies indicates that the gut microbiota plays an important role in cardiovascular health, as reduced microbial diversity has been associated with obesity, diabetes, and coronary artery disease (Kasahara K. et al., 2019). Type2 diabetes (T2D) is well recognized as a major risk factor for the development of cardiovascular disease (CVD), and suggests that alterations in the gut microbiome may partly mediate this association. Specific microbial patterns appear to correlate with early cardiac structural and functional changes in individuals with T2DM. Higher levels of Bacteroidetes, Firmicutes, and Bacteroides are linked with better systolic function, while reduced Firmicutes is associated with left ventricular hypertrophy. Increased Bacteroidetes and Bacteroides relate to fewer diastolic abnormalities, whereas a higher Firmicutes/Bacteroidetes ratio and low Bacteroides levels correspond with enlarged left atrial size. Overall, these microbial patterns suggest that gut microbiome composition may serve as an early marker of subclinical cardiovascular disease in T2D patients (Hui-Ju Tsai et al., 2021). Gut microbiota dysbiosis has been linked to several diseases, including type 2 diabetes and cardiovascular conditions, because it can change the balance of gut bacteria, weaken the intestinal barrier, and increase the release of harmful endotoxins. In patients with heart failure, levels of the phylum Bacteroidetes and the genus Bacteroides are often reduced. Studies also show that people with CVD or heart failure tend to have lower amounts of the beneficial bacterium Faecalibacterium prausnitzii and higher levels of Enterobacteriaceae. These changes suggest that an imbalanced gut microbiome may play an important role in cardiovascular health (Martin kummen et al., 2018).Low levels of Bacteroidetes are linked with both systolic and diastolic dysfunction, while reduced Firmicutes increases the risk of LVH. Studies also show that patients with coronary artery disease or heart failure have lower amounts of Bacteroides. Beneficial metabolites like butyrate and propionate help regulate blood pressure and vascular tone, and butyrate further protects the gut by reducing inflammation and strengthening the intestinal barrier (Melana Yuzefpolskaya et al., 2018). Acetate has been reported to provide protective benefits to the heart by influencing genes involved in cardiac fibrosis and hypertrophy. When short-chain fatty acid (SCFA) producing gut bacteria such as those within the phyla Firmicutes and Bacteroidetes, including the genus Bacteroides are reduced, the resulting decline in SCFA levels may worsen inflammation and more likely to exhibit early abnormalities in left ventricular structure, such as left atrial enlargement and reduced diastolic function (Francine Z. M. et al., 2017). Studies have explored the role of specific microbial metabolites, such as trimethylamine-N-oxide (TMAO), in promoting atherosclerosis and thrombosis. However, few studies have integrated dietary patterns with microbiota diversity to assess their combined impact on CVD risk. Reported that lower adherence to a microbiota-supportive Mediterranean diet was associated with increased visceral adiposity and adverse anthropometric indicators (Kondapalli N et al., 2025). The link between diet-influenced gut microbiota (DI-GM) and cardiovascular disease (CVD) is largely mediated by the gut-heart axis. Diets high in fiber and plant-based foods promote beneficial gut bacteria that produce short-chain fatty acids (SCFAs) like butyrate, which reduce inflammation and help prevent atherosclerosis. Such diets also improve metabolic health by enhancing glucose regulation, balancing lipid levels, and lowering blood pressure, all of which protect the heart (Almeida C et al., 2023).
7. Advanced technology for studying gut microbes:
16S rRNA gene sequencing (Amplicon sequencing):
Sample Collection: Fecal samples are typically collected from participants using sterile, pre-labelled stool collection kits, such as OMNIgene® OMR-200, which stabilize and preserve microbial DNA at ambient temperature for gut microbiome analysis. Participants are provided with standardized instructions and training to ensure proper handling and minimize contamination. Collected samples can be stored at room temperature (approximately 20°C) for short periods, such as 48 hours to 7 days, before being transferred to −80?°C for long-term storage, in accordance with the manufacturer’s recommendations. During transport to the central sequencing facility, ambient temperature is maintained to preserve sample integrity. All samples are processed for genomic DNA extraction within two days of collection, ensuring reliable downstream analysis of the gut microbiome (Raoull Hoogendijk et al., 2024).
DNA Isolation: Bacterial genomic DNA is typically isolated from fecal samples using commercial stool DNA extraction kits, such as the QIAamp DNA Stool Mini Kit (QIAGEN, Hilden, Germany). In general, fecal samples are homogenized in a lysis buffer, and mechanical disruption using beads or a bead?beating device may be applied to improve the breakdown of bacterial cells. Heat treatment is often included to ensure complete lysis. DNA-damaging substances and PCR inhibitors present in stool are removed using inhibitor?removal steps provided in the kit. Purified DNA is eluted in buffer and its quality and concentration are commonly assessed by agarose gel electrophoresis and fluorometric quantification (e.g., Qubit dsDNA HS Assay), producing DNA suitable for downstream microbiome analyses such as 16S rRNA gene sequencing or metagenomics studies (Mirsepasi Het al., 2014).
16S Primers and Amplicon Library Generation:
The hypervariable regions of the 16S rRNA gene, such as V3 and V4, were PCR amplified using extracted microbial DNA as the template, with primer pairs including Probio Uni/Probio Rev for the V3 region. PCR was performed using a suitable DNA polymerase under standard cycling conditions: initial denaturation at 94?°C for 5 minutes, followed by repeated cycles of denaturation at 94?°C for 30 seconds, annealing at 55?°C for 30 seconds, and extension at 72?°C for 90 seconds, with a final extension at 72?°C for 10 minutes. Amplicons were purified using AMPure XP beads, quantified with the Qubit dsDNA HS Assay, normalized to a defined concentration, and pooled in equimolar ratios to generate a sequencing-ready library for downstream high-throughput sequencing (Mancabelli L et al., 2020).
Template Preparation and Sequencing:
When planning sequencing, researchers consider read type, length, depth, and quality. Single-end reads are sequenced in one direction, while paired-end reads, like on Illumina, are sequenced in both directions for longer, more accurate reads. Read length determines which gene regions can be analyzed, and full-length genes require third-generation sequencing. Target regions are PCR-amplified, purified, quantified, and normalized to create an amplicon library. On platforms like Ion Torrent, clonal amplification generates Ion Sphere Particles loaded onto chips for sequencing. Raw reads are cleaned to remove adapters and low-quality sequences, producing high-quality FASTQ data, with ~10,000 reads per sample recommended for complex microbiomes (Watts GS et al., 2017).
Pre-processing Sequence Data:
Sequence data were pre-processed by trimming primers, and reads shorter than 100 bp were discarded. Sequences were then clustered, and those with fewer than 10 copies were filtered out prior to downstream analysis.
Organism Screening and Assessment:
Sequencing reads are mapped to reference 16S rRNA databases, such as the MicroSEQ® 16S rRNA and curated Greengenes databases, using Megablast with an E-value threshold of 0.01 and up to 100 target hits per query. Taxonomic classification requires a minimum of 90% sequence alignment, with genus-level identification assigned at ≥97% identity and species-level at ≥99% identity. Reads that match multiple taxa are evaluated by comparing the top hit to the next best hit; if the difference in sequence identity exceeds 0.2%, the read is assigned to the top hit, otherwise all closely matching taxa within the 0.2% range are reported using a slash ID to indicate ambiguity. Taxonomic abundances are calculated from clustered reads, normalized to relative abundance, and compiled into an OTU table using QIIME workflows optimized for the sequencing platform (Johnson JS et al., 2019).The 16S rRNA gene is a widely used molecular marker for studying bacterial and archaeal phylogeny and taxonomy. It consists of alternating conserved and variable regions: ten conserved regions (C1–C10), which are similar across microorganisms and serve as ideal sites for primer design, and nine hypervariable regions (V1–V9), which accumulate mutations over time and provide species-specific information for microbial identification and classification. While other genes, such as rpoB, can also be used for phylogenetic studies, the 16S rRNA gene is preferred because it is universally present in bacteria and archaea, combines conserved and variable regions suitable for analysis, is long enough to support robust bioinformatic comparisons, retains conserved secondary structures for accurate sequence alignment, and can be easily sequenced, enabling the development of extensive reference databases (Regueira-Iglesias et al., 2023).
Sequence data analysis and statistics:
Sequence reads were quality-filtered, trimmed, and chimeras removed using Mothur (v1.34.1) and UCHIME. Clean reads were clustered into OTUs at 97% similarity and assigned taxonomy using the SILVA or RDP database. Alpha and beta diversity were calculated using the vegan R package, with rarefaction and dendrograms generated for community comparisons, and sequences were normalized across samples. Statistical differences were assessed using AMOVA, parsimony tests, Mann-Whitney U tests with effect sizes, linear regression for environmental trends, and PERMANOVA for community composition. Core microbiota and indicator OTUs (IV ≥ 70, P ≤ 0.05) were identified, and differential taxa determined using Metastats. Community composition and relative abundances were visualized using METAGENassist, MEGAN5, and ggplot2, while maps were produced with ggOceanMaps, ArcGIS, and MATLAB. Data visualization was adapted to sample size, plotting individual points when observations were ≤5 and boxplots for larger groups, with significance set at α = 0.05 (Pascoal F et al., 2024).
Data records: The LogMPIE (Longitudinal Microbiome Profile in Indian Environment) study repository provides three types of data to support microbial analyses.
Data type 1 consists of 1004 FASTQ files from V3 and V4 16S rRNA sequencing of individual subjects, available at the European Nucleotide Archive under accession PRJEB25642/ERP07577, enabling users to perform customized analyses.
Data type 2 includes an OTU table reporting species-level relative abundances across subjects, provided as Supplementary Information.
Data type 3 comprises study metadata, including participant codes, age, sex, BMI, physical activity, and geographic location, facilitating further ecological or epidemiological analyses (Ashok Kumar Dubey et al., 2018).
CONCLUSION:
This review shows that the gut microbiota has a major influence on how type 2 diabetes (T2DM) develops, progresses, and leads to complications. When the gut microbiota becomes unbalanced called gut dysbiosis there is a loss of beneficial short-chain fatty acid (SCFA)–producing bacteria and an increase in harmful, inflammatory microbes. These changes lead to insulin resistance, chronic inflammation, metabolic endotoxemia, and poor glucose control. Microbial metabolites such as SCFAs, bile acids, branched-chain amino acids, imidazole propionate, and lipopolysaccharides (LPS) further affect key metabolic pathways that regulate blood sugar and energy balance.Gut dysbiosis is also linked to serious diabetic complications such as nephropathy, neuropathy, retinopathy, and cardiovascular disease. Each complication shows specific microbial patterns that increase inflammation, oxidative stress, and vascular and immune dysfunction. Diet, lifestyle changes, probiotics, prebiotics, polyphenols, and herbal medicines can help restore healthy gut microbiota and improve metabolic outcomes. Herbal formulations that act through FXR/TGR5 signaling also show promise for better glucose and lipid regulation. New technologies like 16S rRNA sequencing and metagenomics are improving our ability to study gut microbes and understand their role in T2DM. Overall, modifying the gut microbiota offers a promising approach for preventing, detecting, and treating T2DM and its complications. Further research is needed to clarify mechanisms and develop personalized, microbiota-based therapies.
REFERENCES




Dr. Shivkumar S. Ladde*, Madhuri Shinde, Saba Shaikh, Rushikesh Choudhari, Gut Microbiota and Type 2 Diabetes Mellitus: A Review, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 1, 2232-2254. https://doi.org/10.5281/zenodo.18333790
 10.5281/zenodo.18333790
10.5281/zenodo.18333790
