NRI College of Pharmacy, Pothavarappadu, Agiripalli, Eluru
Brain-Derived Neurotrophic Factor (BDNF) is a critical neurotrophin that regulates neuronal survival, differentiation, synaptic plasticity, and cognitive functions. It plays a vital role in both central and peripheral nervous system development, and its dysregulation has been implicated in several neurological and psychiatric disorders, including Alzheimer’s disease, depression, and Parkinson’s disease. BDNF exists in multiple forms preproBDNF, proBDNF, and mature BDNF each exerting distinct biological effects via receptors such as TrkB and p75^NTR. In vitro models offer a controlled environment to study BDNF’s synthesis, function, and response to pharmacological stimuli. However, accurate quantification of BDNF in these systems remains technically challenging due to its low expression levels, structural complexity, and susceptibility to degradation. This review discusses the biological roles and production of BDNF, highlighting the intracellular processing and regulated secretion pathways. It provides a comprehensive overview of in vitro methodologies commonly employed to detect and measure BDNF including ELISA, Western blotting, immunocytochemistry, qRT-PCR, receptor-binding assays, and reporter gene assays while addressing their relative strengths and limitations. Technical challenges such as isoform specificity, cross-reactivity, and assay sensitivity are critically examined. Advances in detection technologies and model systems are also considered. A deeper understanding of these methodological nuances is essential for accurately interpreting BDNF-related findings and for translating in vitro results into clinically relevant insights.
Brain-Derived Neurotrophic Factor (BDNF) is a key protein in the nervous system that plays a vital role in neuronal survival, growth, and plasticity. Belonging to the neurotrophins family, BDNF is extensively expressed in several regions of the brain, particularly in the hippocampus, cortex, and basal forebrain areas that are crucial for higher cognitive functions such as memory formation, learning, and emotional regulation. In both the developing and adult brain, BDNF exerts trophic effects by promoting neuronal differentiation, dendritic arborization, axonal guidance, and synaptic maturation.
Beyond its developmental functions, BDNF remains indispensable in adult neurophysiology, where it regulates synaptic transmission and modulates long-term potentiation (LTP), a cellular correlate of learning and memory. Its actions are mediated predominantly through the tropomyosin receptor kinase B (TrkB), which activates multiple downstream signalling cascades, including the PI3K/Akt, Ras/MAPK, and PLC-γ pathways. These cascades are involved in cellular survival, synaptic efficacy, and metabolic regulation. Notably, BDNF influences both pre- and post-synaptic compartments by enhancing neurotransmitter release and promoting receptor sensitivity, underscoring its critical role in activity-dependent synaptic plasticity.
Importantly, BDNF also exists in two biologically active forms: proBDNF (the precursor) and mature BDNF. These isoforms can exert opposing physiological effects. While mature BDNF predominantly binds to TrkB and promotes cell survival and synaptic strengthening, proBDNF binds to the p75 neurotrophins receptor (p75^NTR^), inducing apoptosis and synaptic weakening. This duality adds a layer of complexity to BDNF signalling and necessitates the development of highly specific and sensitive analytical techniques to discriminate between these forms during quantification.[1]
The dysregulation of BDNF expression and signalling has been implicated in a wide spectrum of neurological and psychiatric disorders. Reduced BDNF levels have been observed in neurodegenerative diseases such as Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, and multiple sclerosis. Furthermore, BDNF has been associated with psychiatric conditions including depression, schizophrenia, and bipolar disorder, where its reduced expression is thought to contribute to impaired synaptic function and mood dysregulation. In animal models and clinical trials, restoring BDNF levels has shown promise in reversing cognitive and behavioural deficits, highlighting its potential as both a biomarker and therapeutic target.
Apart from its role in the central nervous system, BDNF is also involved in the regulation of systemic metabolic processes. Evidence suggests that BDNF influences energy balance by acting on the hypothalamus to suppress appetite and increase energy expenditure. Peripheral or intracerebroventricular administration of BDNF in animal models has led to significant reductions in body weight and food intake, indicating its involvement in obesity and metabolic syndrome.
To advance our understanding of BDNF biology and to develop targeted therapies, in vitro systems such as primary neuronal cultures, neuroblastoma cell lines, stem cell-derived neurons, and organotypic brain slices have become indispensable tools. These systems provide a controlled and reproducible environment to investigate BDNF expression, secretion, and functional responses to pharmacological or genetic manipulations. They allow for the dissection of intracellular signalling mechanisms, study of receptor-ligand interactions, and assessment of neurotrophic responses under various experimental conditions, including oxidative stress, excitotoxicity, or drug treatment.
Despite the utility of in vitro models, accurate and reproducible quantification of BDNF remains a significant technical challenge. The protein is secreted in minute quantities, is sensitive to enzymatic degradation, and is often adsorbed to plastic surfaces in culture systems. Furthermore, the dynamic regulation of BDNF secretion affected by electrical activity, calcium influx, or synaptic stimulation introduces variability that complicates interpretation. Moreover, the presence of multiple BDNF isoforms and post-translational modifications necessitates analytical techniques that can distinguish between biologically distinct forms.[2]
Several methodologies have been employed to quantify BDNF levels in vitro, each with unique strengths and limitations. Enzyme-linked immunosorbent assays (ELISA) remain the most used due to their sensitivity and relative ease of use. However, ELISA may lack specificity for distinguishing proBDNF from mature BDNF unless form-specific antibodies are used. Western blotting can provide information about isoform size and relative abundance but is less quantitative and more time-consuming. Quantitative PCR (qPCR) enables analysis of BDNF mRNA expression, offering indirect insight into BDNF synthesis but not secretion or protein function. More advanced technologies, such as mass spectrometry, offer the promise of highly specific, multiplexed, and isoform-resolved protein quantification but require expensive equipment and technical expertise.
In recent years, novel biosensor-based assays, microfluidic devices, and high-throughput screening platforms have also been developed to improve sensitivity and spatial resolution in BDNF detection. Nonetheless, variability in assay performance, batch-to-batch reagent differences, and lack of standardization remain persistent issues across laboratories. Given the importance of BDNF in neural health and disease, and the technical intricacies associated with its measurement, it is critical to evaluate the methodologies available for its quantification in in vitro systems. This review aims to (1) summarize the existing approaches for measuring BDNF in cultured cell and tissue models, (2) critically assess their advantages and limitations, and (3) highlight emerging technologies and best practices that may improve accuracy, reproducibility, and biological relevance of BDNF quantification. Through this discussion, we aim to provide a practical and up-to-date resource for researchers seeking to unravel the complexities of BDNF signalling in experimental neuroscience.[3]
Table 1: Comparitive Overview of BDNF Quantification Methods in In Vitro Systems.
|
Method |
Principle |
Advantages |
Limitations |
Detects proBDNF vs. mature BDNF? |
|
ELISA (Enzyme-Linked Immunosorbent Assay) |
Antibody-based detection of BDNF in culture supernatants or lysates |
- High sensitivity- Quantitative- Suitable for high-throughput screening |
- May not differentiate between proBDNF and mature BDNF unless specific antibodies are used- Batch variability in kits |
Only if isoform-specific antibodies are used |
|
Western Blotting |
Separation by electrophoresis and detection via antibodies |
- Allows size-based discrimination (proBDNF vs. mature BDNF)- Semi-quantitative- Confirms protein integrity |
- Low throughput- Labor-intensive- Requires large amounts of sample- Less sensitive than ELISA |
Yes |
|
qPCR (Quantitative Polymerase Chain Reaction) |
Measures BDNF mRNA expression |
- Sensitive- Quantitative- Indicates gene expression regulation |
- Does not measure protein or secreted BDNF- mRNA levels may not correlate with protein abundance |
No (RNA level only) |
|
Mass Spectrometry (MS) |
Detects and quantifies peptides based on mass/charge |
- High specificity- Can distinguish isoforms and post-translational modifications- Multiplexing possible |
- Expensive- Requires technical expertise- Low throughput |
Yes |
|
Luminex Bead-Based Assay |
Multiplex immunoassay using fluorescent beads |
- Detects multiple analytes simultaneously- High sensitivity- small sample volume needed |
- Requires specialized equipment- Limited isoform resolution |
Rarely (depends on antibody selection) |
|
ELISPOT (Enzyme-Linked ImmunoSpot) |
Detects single cells secreting BDNF |
- High resolution at single-cell level- Useful for studying secretion dynamics |
- Semi-quantitative- Complex setup- May require optimization |
No |
|
Bioassay (e.g., TrkB activation) |
Functional assays using cells expressing TrkB to detect BDNF activity |
- Biologically relevant- Detects functional (active) BDNF |
- Indirect- No isoform distinction- Less quantitative |
No |
|
Immunocytochemistry / Immunofluorescence |
Visualizes BDNF localization within cells |
- Spatial resolution- Allows co-localization studies |
- Qualitative or semi-quantitative- Subjective analysis- Not ideal for soluble BDNF |
Rarely |
Structure of BDNF:
BDNF has close structural homology to NGF and shares about 50% amino acid identity with NGF, NT-3 and NT-4/5. Each neurotrophin consists of a non-covalently linked homodimer with a signal peptide following the initiation codon and pro-region containing an N-linked glycosylation site. In rats, the BDNF gene is located on chromosome 11 and is controlled by multiple activity-dependent and tissue-specific promoters I, II, III, IV; cAMP response-element binding protein (CREB) and upstream stimulatory factor-1/2 (USF-1/2) regulate promoters I and III, and calcium responsive transcription factor (CaRF) mediates transcription by binding to promoter III. All exons that have been defined in humans are also expressed in mouse and rat, except for human exons VIIB and VIII. The rat BDNF gene has been suggested to undergo cryptic splicing within exon II to form IIA, IIB and IIC genes . The mouse BDNF gene has eight exons containing separate promoters upstream of each exon and one 3’ exon encodes the mature BDNF protein. Multiple promoters determine tissue-specific expression of the BDNF transcript. Human BDNF structure is closely related to rat and mouse BDNF . Eight distinct mRNAs are transcribed, with transcripts containing exons I–III expressed predominantly in brain and exon IV found in lung and heart. In situ hybridization experiments have revealed that BDNF mRNA is strongly expressed in the brain. The BDNF expression levels are low during foetal development, markedly increase after birth, and then decrease in adults.[4]
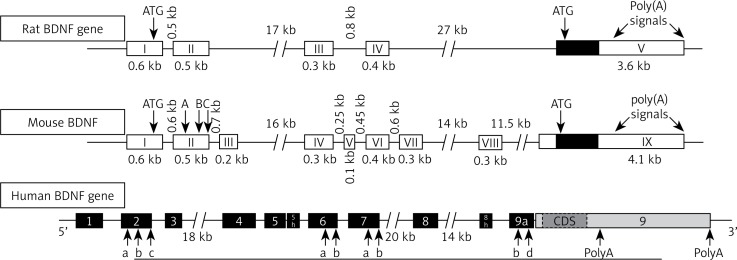
Figure 1: Gene structure of BDNF.
Biology of BDNF:
Brain-Derived Neurotrophic Factor (BDNF) is one of the most extensively studied neurotrophins due to its essential role in neuronal survival, development, synaptic plasticity, and cognitive processes such as learning and memory. Since its discovery in 1982, BDNF has been recognized as a key signalling molecule within the central and peripheral nervous systems. The synthesis and maturation of BDNF involve a tightly regulated, multi-stage process that includes the generation of several precursor forms, intracellular trafficking, and secretion, each contributing to its diverse and context-dependent biological functions.
1. BDNF Gene and mRNA Regulation
The human BDNF gene is located on chromosome 11 (11p14.1) and is composed of multiple exons and alternative promoters. This complex structure allows the production of various BDNF transcript variants through alternative splicing and promoter usage. While all transcripts encode the same BDNF protein, their different 5' untranslated regions (UTRs) confer spatial and temporal control of expression. Activity-dependent expression of BDNF, especially in neurons, is regulated by several signalling cascades—including calcium influx, cAMP-response element-binding protein (CREB) activation, and mitogen-activated protein kinase (MAPK) pathways. These mechanisms ensure that BDNF production is tightly linked to neuronal activity, making it a crucial mediator of synaptic modulation and long-term potentiation (LTP).
2. Protein Structure and Conservation
The BDNF protein is highly conserved across species, consisting of 247 amino acids in humans. It belongs to the neurotrophins family, which includes nerve growth factor (NGF), neurotrophin-3 (NT-3), and neurotrophin-4/5 (NT-4/5). The mature BDNF structure forms a homodimer and exhibits the characteristic "cysteine knot" motif—a compact structural arrangement stabilized by three disulfide bonds critical for receptor binding. The conservation of its sequence and structure underscores its fundamental biological importance in neurodevelopment and synaptic function.
3. Multistage Processing: PreproBDNF to Mature BDNF
BDNF biosynthesis occurs via a series of post-translational processing steps that begin in the endoplasmic reticulum (ER) and proceed through the Golgi apparatus and secretory vesicles.
a. PreproBDNF (32–35 kDa)
The initial translation product, preproBDNF, contains a signal peptide ("pre" region) that directs the nascent polypeptide into the rough ER. Within the ER, the signal sequence is rapidly cleaved, yielding the proBDNF intermediate. The prepro form is not biologically active but is essential for proper targeting and folding of the protein.
b. ProBDNF (28–32 kDa)
The proBDNF form contains an N-terminal pro-domain and a C-terminal mature domain. While initially considered an inactive precursor, proBDNF is now recognized to have distinct biological activity, often mediating effects opposite to those of mature BDNF. ProBDNF preferentially binds to the p75 neurotrophin receptor (p75^NTR) and co-receptors like sortilin, activating apoptotic or growth-inhibitory signalling cascades. This highlights the importance of regulated cleavage to maintain a balance between neurotrophic support and pruning or death.
c. Mature BDNF (13–14 kDa)
The final bioactive product, mature BDNF (mBDNF), is produced via proteolytic cleavage of proBDNF. This cleavage can occur intracellularly or extracellularly, depending on the cellular context and availability of specific proteases. mBDNF primarily binds to the tropomyosin-related kinase B (TrkB) receptor, initiating a cascade of signalling pathways (e.g., PI3K/Akt, MAPK/ERK, PLCγ) that promote neuronal survival, differentiation, synaptic strengthening, and plasticity.
4. Proteolytic Cleavage Mechanisms
The conversion of proBDNF to mature BDNF is mediated by several proteolytic systems:
Inside the trans-Golgi network or in secretory vesicles, proconvertases from the subtilisin/kexin-like family (e.g., furin, PACE4, and PC1/3) cleave proBDNF to produce mBDNF. These enzymes recognize specific consensus motifs within the pro-domain and are active in acidic environments typically found in secretory granules.
In some neurons, proBDNF is secreted into the extracellular space, where it can be processed by metalloproteinases (e.g., MMP-2, MMP-9) or plasmin, a serine protease activated from plasminogen by tissue-type plasminogen activator (tPA). This step is crucial for synaptic remodelling and activity-dependent plasticity, allowing fine-tuning of BDNF function based on extracellular cues.
The precise regulation of when and where this cleavage occurs is vital for maintaining neuronal homeostasis. Dysregulation in this process is implicated in several neurological and psychiatric disorders, including depression, Alzheimer’s disease, and schizophrenia.
5. Intracellular Trafficking and Secretion
After processing in the Golgi, BDNF is packaged into secretory vesicles, which are then transported to axon terminals or dendritic compartments. The release of BDNF is highly activity-dependent.
Some cells secrete BDNF continuously at low levels, contributing to baseline trophic support.
In neurons, depolarization or calcium influx triggers vesicular fusion with the plasma membrane, releasing BDNF into the synaptic cleft. This activity-dependent secretion is critical for synaptic modulation, LTP induction, and dendritic spine remodelling.
Interestingly, BDNF can also be retrogradely transported, particularly in developing neurons. When secreted from target tissues, BDNF can bind to TrkB receptors at axon terminals and be internalized into signalling endosomes, which are transported back to the soma to influence gene transcription and cell fate.
6. Functional Divergence of BDNF Isoforms
The proBDNF/mBDNF ratio is therefore a critical determinant of cellular outcomes and is modulated in response to neuronal activity, stress, and disease states.[5]
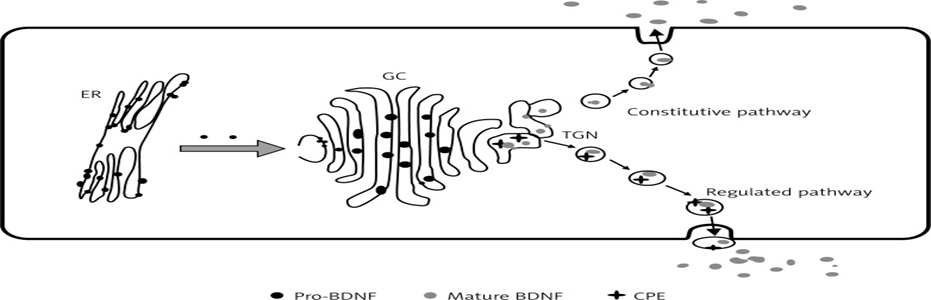
Figure 2: origin of BDNF.
Production of BDNF:
Brain-Derived Neurotrophic Factor (BDNF) production is a tightly regulated biological process that involves transcriptional activation, mRNA splicing, translation, post-translational modifications, and secretion. This multilayered regulation allows BDNF to respond dynamically to physiological demands such as neuronal activity, development, injury, and environmental stimuli like exercise or stress.
1. Gene Structure and Transcription
The BDNF gene is located on chromosome 11p14.1 in humans and is composed of multiple 5′ non-coding exons and a single 3′ coding exon. These non-coding exons are each under the control of distinct promoters, leading to multiple mRNA variants through alternative splicing. All variants ultimately encode the same BDNF protein but allow spatially and temporally distinct expression profiles in different brain regions or developmental stages.
2. mRNA Transport and Localization
After transcription, BDNF mRNA is exported from the nucleus and, in neurons, may be transported to dendrites or axons. This spatial localization is important for localized protein synthesis at synapses in response to stimuli, contributing to synaptic plasticity. RNA-binding proteins and cytoskeletal transport systems mediate this trafficking.
3. Translation and Precursor Formation
The translation of BDNF mRNA in the cytoplasm produces a preproBDNF protein (32–35 kDa), which includes:
This precursor is directed into the rough endoplasmic reticulum (ER), where it undergoes initial folding and processing. The signal peptide is cleaved, producing proBDNF (28–32 kDa). Proper folding is assisted by chaperone proteins in the ER.
4. Post-Translational Modifications and Maturation
The proBDNF is transported to the Golgi apparatus and subsequently into secretory vesicles, where it can undergo further processing. There are two main fates for proBDNF:
This processing is biologically significant as:
5. Secretion Pathways
BDNF is secreted from cells via two major mechanisms:
Calcium-triggered exocytosis ensures that BDNF release is synchronized with synaptic activity, enabling its function in synaptic plasticity and long-term potentiation (LTP).
6. Factors Influencing BDNF Production
Several internal and external stimuli influence BDNF gene expression and protein production:
Table 2: Several internal and external stimuli influence BDNF gene expression and protein production
|
Stimulus |
Effect on BDNF Production |
|
Neuronal activity |
↑ BDNF transcription via calcium influx and CREB activation |
|
Physical exercise |
↑ BDNF levels in brain and serum |
|
Environmental enrichment |
↑ BDNF expression in hippocampus and cortex |
|
Stress |
↓ BDNF levels (especially chronic stress and cortisol exposure) |
|
Antidepressants |
↑ BDNF transcription (mechanism of SSRI efficacy) |
|
Neurodegenerative diseases |
↓ BDNF expression in Alzheimer's, Parkinson's, and Huntington's |
|
Hypoxia or ischemia |
Can trigger compensatory BDNF upregulation for neuroprotection |
7. In Vitro Considerations
In cultured cells, BDNF production can be induced or modulated by various stimuli:
However, many in vitro systems have inherently low BDNF expression and may require optimization of conditions, cell types, and detection methods to reliably measure production.[6]
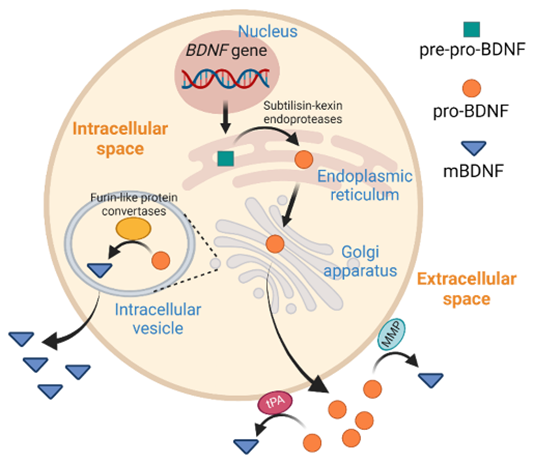
Figure 3: Production of BDNF.
Plasma levels of BDNF:
In healthy volunteers, mean plasma BDNF level was found to be ∼92.5 pg/ml (8.0–927.0 pg/ml). It was higher in women, and decreased with advancing age in both genders . BDNF is widely distributed in various regions of the brain, and aids in survival, support, and function of neurons. Other sources of BDNF include the lungs, heart, spleen, gastrointestinal tract, and liver. Apart from these, BDNF was found to be expressed in fibroblasts, vascular smooth muscle cells, and thymic stroma . Studies of Lommatzsch et al. confirmed that levels of BDNF in the urinary bladder, lung, and colon were higher than those found in the brain or skin. A positive correlation between blood levels of BDNF and diastolic blood pressure, total cholesterol, low-density lipoprotein (LDL) cholesterol, adipose tissue mass, body mass index and triglyceride were reported. It was suggested that women with low plasma BDNF levels have increased mortality risk. This is supported by the observation that a significant reduction in plasma levels of BDNF in females correlated with advancing age and body weight.[7]
Agonism of BDNF action:
BDNF mRNA expression is regulated by neuronal activity since epileptogenic activation of glutamatergic synapses increased the expression of BDNF mRNA in slices of rat hippocampus. Increase in synaptic activity induced by AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) receptor agonists produced a transient elevation in mRNA levels encoding BDNF and TrkB in the hippocampus and cortex. Light stimulation increased BDNF mRNA in the visual cortex, osmotic stimulation enhanced BDNF mRNA in the hypothalamus, and whisker stimulation increases BDNF mRNA expression in the somatosensory barrel cortex. Electrical stimuli that induce long-term potentiation (LTP) in the hippocampus, a cellular model of learning and memory, increased BDNF and NGF expression. Physical exercise increases NGF and BDNF expression in the hippocampus, which may explain its beneficial action in improving memory and prevent the development of Alzheimer's disease. BDNF interacts with oestrogen to induce neuropeptide-Y (NPY) and together the trio (BDNF-NPY-oestrogen) regulates the function of the hippocampus.
Antagonism of BDNF action:
Blockade of glutamate receptors or stimulation of the GABAergic (gamma-amino butyric acid) system reduced BDNF mRNA level in the hippocampus. Bozzi et al. demonstrated that an increase or a decrease of neuronal activity can enhance or reduce BDNF expression. Consistent with these observations, light-induced physiological activity enhanced the expression of BDNF mRNA in the visual cortex and monocular deprivation elicited a striking decrease in BDNF mRNA in the visual cortex corresponding to the deprived eye. Anti-BDNF antiserum and the TrkB-IgG construct have been successful to antagonize the action of BDNF in neuropathic pain and inflammatory hypersensitivity models. These findings demonstrate the potential for antibody-mediated TrkB agonism as a potential therapeutic approach to enhance retinal ganglion cells’ (RGC) survival after optic nerve injury. Eibl et al. reported that Y1036, a potential multipotent neurotrophin antagonist, altered molecular topology (surface charge density) and acted against BDNF and NGF by preventing NT-induced receptor activation and downstream signalling via the p44/42 MAPK pathway. Furthermore, endogenous production of nitric oxide (NO) downregulated BDNF secretion in hippocampal neurons by activating a cyclic guanosine monophosphate (cGMP)-dependent signal transduction pathway leading to downregulation of Ca2+ release from IP3-sensitive intracellular stores by protein kinase G.[8,9]
Signalling cascade in BDNF:
BDNF (ligand) activation of tyrosine residues results in activation of different intracellular pathways, as shown in (figure 4), leading to neural plasticity, neurogenesis, stress resistance and survival of the cell. This suggests comparative flexibility of Trk receptors in terms of pro-survival function. Thus, BDNF signalling pathways activate one or both transcription factors CREB and CREB-binding protein (CBP) that regulate expression of genes encoding proteins involved in neural plasticity, stress resistance and cell survival.[10]

Figure 4: Signalling Pathway of BDNF.
Functions of Brain-Derived Neurotrophic Factor (BDNF):
Brain-Derived Neurotrophic Factor (BDNF) plays a central role in the development, maintenance, and plasticity of the nervous system. As a member of the neurotrophin family, BDNF exerts a wide array of biological functions through its interactions with high-affinity TrkB receptors and the low-affinity p75 neurotrophin receptor (p75^NTR). These interactions trigger multiple intracellular signalling pathways that modulate neuronal physiology and behaviour. The following section outlines the major functions of BDNF in both the central and peripheral nervous systems.
1. Neuronal Survival and Differentiation
BDNF is essential for the survival of sensory neurons, motor neurons, and various subtypes of central neurons. During development, BDNF supports neuronal differentiation and maturation, particularly in the hippocampus, cortex, and basal forebrain. It promotes axonal and dendritic growth, enhances arborization, and prevents apoptosis by activating pro-survival signalling pathways such as PI3K/Akt. These effects are critical for establishing and refining neural circuits.
2. Synaptic Plasticity and Long-Term Potentiation (LTP)
One of the most widely studied functions of BDNF is its role in modulating synaptic plasticity, particularly long-term potentiation (LTP) a cellular mechanism underlying learning and memory. BDNF enhances synaptic efficacy by increasing neurotransmitter release at presynaptic terminals and upregulating receptor sensitivity and density at postsynaptic sites. These actions are especially pronounced in hippocampal neurons, where BDNF-TrkB signalling facilitates both structural and functional synaptic changes required for memory consolidation.
3. Neurogenesis
BDNF plays a crucial role in adult neurogenesis, particularly in the sub granular zone (SGZ) of the dentate gyrus in the hippocampus. It promotes the proliferation, differentiation, and survival of neural progenitor cells and facilitates their integration into existing neural networks. This process is critical not only for cognitive adaptability but also for emotional regulation and recovery from brain injury.
4. Regulation of Intracellular Signalling Pathways
Upon binding to the TrkB receptor, BDNF activates several downstream intracellular signalling cascades, including:
These pathways regulate a variety of cellular processes including gene transcription, cytoskeletal dynamics, and vesicle trafficking.
5. Modulation of Gene Expression
BDNF influences gene expression by activating transcription factors such as CREB (cAMP response element-binding protein), which regulate the expression of activity-dependent genes important for synaptic plasticity. This transcriptional regulation supports long-term synaptic changes and memory formation.
6. Cognitive and Behavioural Functions
BDNF has a significant impact on cognitive function, particularly learning and memory. Experimental studies have shown that reduced BDNF levels are associated with impaired spatial learning and memory retention. Moreover, BDNF plays a role in emotional regulation, and reduced expression has been implicated in the pathophysiology of mood disorders such as depression and anxiety. The ability of antidepressants and physical activity to increase BDNF levels further underscores its importance in behavioural regulation.
7. Neuroprotection
BDNF exhibits potent neuroprotective properties. It protects neurons from a variety of insults, including glutamate-induced excitotoxicity, oxidative stress, hypoxia, and neurotoxic agents. BDNF-mediated activation of TrkB signalling helps maintain mitochondrial function, reduces inflammation, and prevents apoptotic cell death. These neuroprotective functions are especially relevant in conditions like stroke, epilepsy, and neurodegenerative diseases.
8. Regulation of Energy Homeostasis and Metabolism
Emerging research indicates that BDNF also regulates energy balance and metabolism. BDNF expression in the hypothalamus plays a role in appetite suppression and body weight regulation. Experimental administration of BDNF has been shown to decrease food intake and increase energy expenditure. Additionally, BDNF is involved in the regulation of glucose metabolism and insulin sensitivity, linking it to the pathophysiology of obesity and metabolic syndrome.
9. Therapeutic Relevance
Given its multifaceted role, BDNF is increasingly recognized as a promising therapeutic target. Reduced BDNF levels have been consistently observed in various neurological and psychiatric disorders, including Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, depression, schizophrenia, and traumatic brain injury. Efforts to modulate BDNF levels through pharmacological agents, physical activity, or gene therapy are being explored as potential treatments to restore synaptic function and neuronal health.[11]
10. Actions on cardiac and endothelial cells
Neurotrophins promote angiogenesis and control survival of adult endothelial cells (EC), vascular smooth muscle cells (VSMC) and cardiomyocytes. It was reported that BDNF promoted therapeutic neovascularization via the TrkB receptor, whereas low-affinity receptor p75 NTR not only induced apoptosis of endothelial cells and vascular smooth muscles but also impaired angiogenesis. Studies performed in a murine BDNF knockout model revealed that NT-3 and BDNF are involved in the formation of heart and myocardial vasculature. TrkB receptors expressed on endothelial cells activate two major signalling pathways, ERK/MAPK and PI3-kinase/AKT, to promote EC survival. In addition, AKT activated endothelial nitric oxide (NO) synthase, which contributes to vascular relaxation; this may account for its cardiovascular protective action.[12]
11. Role in inflammation and immunity
Elevated levels of neurotrophins (NTs) may contribute to the development of bronchial hyperreactivity (BHR), a hallmark of allergic asthma, which is confirmed from the observation that release of NTs occurred from immune cells including B-lymphocytes, eosinophils, mast cells and macrophages. It is likely that upon stimulation with antigen (Ag), CD4+ T cells produce BDNF by truncated gp95TrkB (expressed in non-neuronal tissues). BDNF may act as a mediator between airway inflammatory events and neuronal changes that occur during the induction of allergic asthma by enhancing airway smooth muscle contraction and mucus hypersecretion by facilitating the release of acetyl choline and plasma extravasation (Figure 5). On the other hand, in neuro-inflammatory diseases such as multiple sclerosis, there may occur enhanced production of BDNF, which may show neuro-protective activity due to its immunomodulatory action. Thus, BDNF can be used as a therapeutic strategy in detection and prevention of neurological inflammatory disorders.[13]
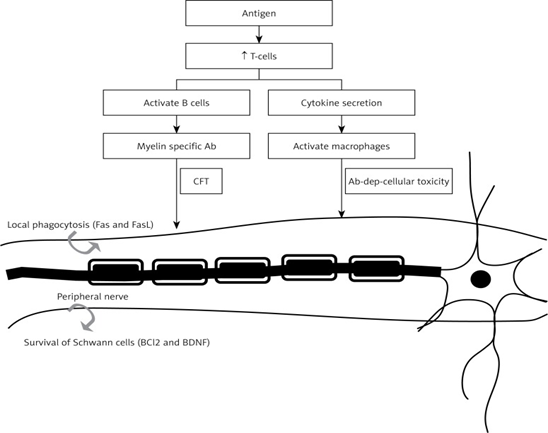
Figure 5 : Mechanism of BDNF in Inflammation and Immunity.
12. Lipid metabolism
In 1995, it was reported that BDNF affects energy metabolism following intracerebroventricular (ICV) administration, which leads to decreased energy intake and body weight loss in rats. This action of BDNF was found to be associated with a dose-dependent increase in serotonin turnover, nerve cell survival and adaptive plasticity. BDNF showed a positive correlation with low-density lipid (LDL) cholesterol, total cholesterol, and triglycerides. BDNF treatment of diabetic animals resulted in a decrease of plasma glucose, non-esterified fat, phospholipids, and liver weight, along with an increase in β-oxidation, peroxisome proliferator activator receptor (PPAR-α) activation and level of fibroblast growth factor. TrkB activation is essential for appetite regulation and energy homeostasis. It was reported that dietary omega-3 fatty acids normalize BDNF levels (Figure 6), reduce oxidative damage and improve learning ability after traumatic brain injury.[14]
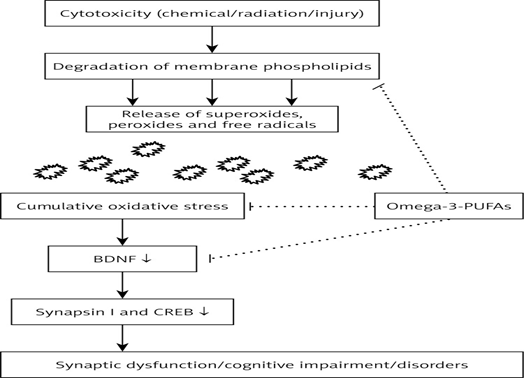
Figure 6: Role of BDNF in Lipid Metabolism.
13. BDNF in type-2 diabetes mellitus:
BDNF has been implicated in the regulation of food intake and body weight both in experimental animals and humans. For instance, systemic administration of BDNF decreased non-fasted blood glucose in obese, non-insulin-dependent diabetic C57BLKS-Lepr(db)/lepr(db) (db/db) mice, with a concomitant decrease in body weight. The effect of BDNF on non-fasted blood glucose levels is not caused by decreased food intake but was found to be due to improvement in blood glucose control, an effect that persisted for weeks after cessation of BDNF treatment. BDNF reduced hepatomegaly present in db/db mice, reduced liver glycogen and reduced liver enzyme activity in serum, supporting the involvement of liver tissue in the mechanism of action of BDNF.
Further support for the role of BDNF in diabetes mellitus is evident from the observation that once or twice per week administration (70 mg/kg/wk) to db/db mice for 3 weeks significantly reduced blood glucose concentrations and hemoglobin A1c (HbA1c) compared to controls. These results suggest that BDNF not only reduced blood glucose concentrations but also restored systemic glucose balance, indicating the possibility that BDNF could be a novel hypoglycemic agent even with treatment as infrequently as once per week. Furthermore, intracerebroventricular (ICV) administration of BDNF lowered blood glucose, increased pancreatic insulin content, enhanced thermogenesis and norepinephrine turnover, and increased uncoupling protein-1 mRNA expression in the interscapular brown adipose tissue of db/db mice. This evidence indicates that BDNF activates the sympathetic nervous system and thus regulates energy expenditure in obese diabetic animals.
The role of BDNF in type 2 diabetes mellitus is further supported by the observation that plasma levels of BDNF were decreased in humans with type 2 diabetes mellitus independently of obesity, and inversely associated with fasting plasma glucose, but not with insulin. When output of BDNF from the human brain was studied, output was inhibited when blood glucose levels were elevated, whereas when plasma insulin was increased while maintaining normal blood glucose, the cerebral output of BDNF was not inhibited. These results indicate that high levels of glucose, but not insulin, inhibited the output of BDNF from the human brain. These results emphasize that low levels of BDNF accompany impaired glucose metabolism, and decreased BDNF may be a factor involved in type 2 diabetes mellitus.[15]
Mechanism(s) involved in the cytoprotective action of BDNF in type 2 diabetes mellitus:
BDNF not only regulated glucose and energy metabolism but also prevented exhaustion of the pancreas in diabetic mice by maintaining the histologic cellular organization of β cells and non-β cells in pancreatic islets and restoring the level of insulin-secreting granules in β cells and thus ameliorated diabetes in experimental animals. These actions of BDNF suggest that it could protect pancreatic β cells. The possible mechanism(s) involved in this cytoprotective action of BDNF could be attributed to its binding to the high-affinity receptor TrkB, resulting in the recruitment of proteins that activate IRS-1/2, PI-3K and protein kinase B (Akt). These actions result in activation of one or more transcription factors (CREB and CBP) that regulate expression of genes encoding proteins involved in neural plasticity, stress resistance and cell survival. Furthermore, BDNF and IGF-I have similar downstream signalling mechanisms, incorporating both p-CAMKII (calcium/calmodulin protein kinase II) and p-MAPKII (phosphorylated mitogen-activated protein kinase II) signalling cascades, which promote cell survival by a dual mechanism consisting of posttranslational modification and inactivation of a component of the cell death machinery and increased transcription of pro-survival genes. In an extension of this study, it was noted that two chemically distinct inhibitors of PI-3K, wortmannin and LY294002, reduced PI-3K activation by IGF-1 and inhibited its survival-promoting activity, suggesting that PI-3K is necessary for IGF-1-mediated survival. Ultimately death resulting from PI 3-kinase blockade is accompanied by DNA fragmentation, a hallmark of apoptosis. It is likely that BDNF may have actions like IGF-1 in producing its cytoprotective actions on pancreatic β cells.[16]

Figure 7: Possible mechanism(s) involved in cytoprotective action of BDNF.
Clinical significance of BDNF:
It is evident from the preceding discussion that BDNF has several important actions that could have significant clinical implications. Decrease in the expression of BDNF is seen in many neurological diseases such as Alzheimer's disease, Parkinson's disease, Huntington's disease, and bipolar disease , as detailed above. Physical exercise enhances brain BDNF levels and thus improves in depression. Lithium, which is used in bipolar disorder, is known to enhance TrkB activation and expression of BDNF mRNA, which supports the role of BDNF in bipolar disorder. Interestingly, overexpression of BDNF in the hippocampus was found to occur in spontaneous seizures, leading to temporal lobe epilepsy. Furthermore, BDNF is synthesized and released in rabbit intestinal smooth muscle cells (SMCs), which is influenced by release of calcium levels that, in turn, activate substance-P (SP) along with pituitary adenylate cyclase activating peptide (PACAP). BDNF can modulate gut function by activating the PLC pathway and hence could have therapeutic potential in the treatment of irritable bowel syndrome and functional dyspepsia. A significant decrease in the levels of BDNF-BCl2-Akt (genes involved in anti-apoptotic signalling pathways of BDNF) was observed in autism disorder. Thus, plasma levels of BDNF may be used as a bio-marker for detection of autism disorder during early stages. As already discussed above, BDNF has an important role in energy homeostasis that accounts for its role in obesity, type 2 diabetes mellitus and metabolic syndrome. BDNF, both by its peripheral and central actions, seems to be capable of preventing type 2 diabetes mellitus. It remains to be seen whether administration of BDNF could constitute a new therapeutic approach in the management of obesity, type 2 diabetes mellitus and metabolic syndrome, in both their prevention and management.[17]
In Vitro Methods Used to Study BDNF Protein:
Brain-Derived Neurotrophic Factor (BDNF) is a crucial neurotrophin involved in neuronal survival, synaptic plasticity, and cognitive functions. Understanding BDNF’s structure, function, and signalling mechanisms requires a range of in vitro experimental approaches. These techniques allow researchers to study BDNF in a controlled environment, independent of the complex variables found in living organisms. Below are the principal in vitro methods used to study BDNF protein, discussed individually.
1. Recombinant Protein Expression
Recombinant DNA technology is widely used to express BDNF protein in various host systems. The BDNF gene is cloned into an appropriate expression vector and introduced into host cells such as Escherichia coli, HEK293, or CHO cells. While bacterial systems offer simplicity and high yield, mammalian expression systems are preferred for BDNF due to their ability to support proper protein folding, glycosylation, and secretion. Induction with agents like IPTG (in bacterial systems) or CMV promoter activity (in mammalian systems) triggers BDNF expression, which is then harvested from the culture medium or cell lysate.
Systems Used:
Applications:

2. Protein Purification Techniques
Following recombinant expression, BDNF protein is purified to isolate it from other cellular components. Affinity chromatography is the most common method, especially when BDNF is expressed with a fusion tag such as a His-tag or FLAG-tag. The lysate is passed through a column containing a resin that binds the tagged BDNF, while impurities are washed away. The protein is then eluted using an imidazole gradient or specific buffer conditions. Purified BDNF is essential for downstream functional and biochemical assays.
Common Methods:
Applications:
3. Western Blotting
Western blot analysis is employed to detect and confirm the presence of BDNF protein in cell lysates or conditioned media. Samples are separated by SDS-PAGE and transferred onto a nitrocellulose or PVDF membrane. The membrane is probed with a primary antibody specific to BDNF, followed by a secondary antibody conjugated to an enzyme (e.g., HRP). Detection is achieved via chemiluminescence or fluorescence. This method not only verifies protein expression but can also distinguish between proBDNF and mature BDNF forms.
Procedure:
Applications:
4. Enzyme-Linked Immunosorbent Assay (ELISA)
ELISA is a quantitative method widely used to measure BDNF concentrations in biological samples. In a typical sandwich ELISA, a capture antibody specific to BDNF is immobilized on a microplate. The sample is added, allowing BDNF to bind the capture antibody. A secondary detection antibody, linked to an enzyme such as horseradish peroxidase (HRP), produces a colorimetric signal upon substrate addition. The intensity of the signal is proportional to the BDNF concentration and is measured using a spectrophotometer.
Procedure:
Applications:
5. Immunocytochemistry (ICC) and Immunofluorescence (IF)
These techniques are utilized to visualize the intracellular and extracellular localization of BDNF within cultured cells. Cells are fixed and incubated with a primary antibody targeting BDNF, followed by a fluorophore-conjugated secondary antibody. The samples are then examined under a fluorescence microscope or confocal microscope. ICC/IF provides spatial information on BDNF expression patterns and helps elucidate its trafficking and secretion dynamics in neuronal cells.
Procedure:
Applications:

6. Quantitative Real-Time PCR (qRT-PCR)
Although not a direct protein assay, qRT-PCR is routinely employed to evaluate BDNF gene expression, serving as an indirect indicator of potential protein production. Total RNA is extracted from cultured cells and reverse transcribed into cDNA. Real-time PCR, using primers specific for BDNF mRNA, allows precise quantification of transcript levels. This method is especially useful when assessing transcriptional regulation in response to various stimuli or pharmacological treatments.
Procedure:
Applications:

7. Neurite Outgrowth Assays
To assess the biological activity of BDNF, neurite outgrowth assays are commonly conducted using neuronal cell lines such as PC12 or primary neurons. These cells are cultured in the presence of recombinant BDNF, and their morphological response such as neurite extension is measured. BDNF stimulates TrkB receptor activation, triggering intracellular signalling cascades that promote neuronal differentiation and neurite elongation. This functional assay is crucial for evaluating the potency of BDNF preparations.
Procedure:
Applications:

8. Receptor Binding Assays
BDNF exerts its effects primarily through binding to the high-affinity TrkB receptor. In vitro binding assays are used to quantify this interaction. Techniques such as radioligand binding, surface plasmon resonance (SPR), or fluorescence-based assays can be employed to study BDNF-TrkB kinetics, affinity, and specificity. These methods are vital for drug screening and for understanding BDNF’s mechanism of action.
Techniques Used:
Applications:

9. Cell-Based Reporter Gene Assays
Reporter assays provide a robust method to measure the downstream signalling activity of BDNF. Cells are transfected with reporter constructs such as luciferase or GFP under the control of promoters responsive to TrkB activation (e.g., CRE, NFAT). Upon BDNF treatment, activation of intracellular pathways leads to transcription of the reporter gene, which can then be quantified. This approach is useful for evaluating functional responses in a high-throughput format.
Procedure:
Applications:
Table 3: Summary of In Vitro Methods.
|
Method |
Purpose |
Key Components / Techniques |
Applications |
|
1. Recombinant Protein Expression |
To produce BDNF in cell systems |
E. coli, HEK293, CHO cells; expression vectors with promoters (e.g., CMV); IPTG or other inducers |
Producing BDNF for functional assays, structural studies, or therapeutic screening |
|
2. Protein Purification |
Isolate BDNF from cell lysates or media |
Affinity chromatography (His-tag, FLAG-tag), size-exclusion, ion-exchange |
Preparation for biochemical assays, receptor binding studies, or animal injections |
|
3. Western Blotting |
Detect and differentiate BDNF protein isoforms |
SDS-PAGE, PVDF/nitrocellulose membrane, BDNF-specific antibodies, chemiluminescent substrates |
Distinguishing proBDNF vs mature BDNF; expression validation; treatment monitoring |
|
4. ELISA |
Quantify BDNF levels in media or lysates |
Capture & detection antibodies, HRP enzyme, colorimetric/fluorescent readout |
Monitoring BDNF secretion; high-throughput quantification; drug screening |
|
5. Immunocytochemistry / Immunofluorescence (ICC/IF) |
Visualize BDNF localization in cells |
Fixed cells, fluorophore-tagged antibodies, confocal or fluorescence microscopy |
Subcellular localization; co-localization with receptors or organelles; trafficking studies |
|
6. qRT-PCR |
Quantify BDNF mRNA expression (indirect protein estimate) |
RNA isolation, reverse transcription, specific primers, SYBR Green/TaqMan probes |
Gene expression profiling; effects of drugs or stimuli on BDNF transcription |
|
7. Neurite Outgrowth Assays |
Evaluate biological activity of BDNF on neurons |
PC12/SH-SY5Y/primary neurons, image analysis software |
Functional validation of recombinant BDNF; neurotrophic potency assays |
|
8. Receptor Binding Assays |
Assess BDNF interaction with TrkB receptor |
Radiolabelled BDNF, SPR, fluorescence-based ligand binding assays |
TrkB binding affinity/specificity; screening agonists/antagonists |
|
9. Reporter Gene Assays |
Measure downstream signalling activity of BDNF |
Luciferase/GFP reporters under control of CRE/NFAT promoters |
Functional pathway analysis; high-throughput drug screening; real-time BDNF activity measurement |
Technical challenges:
Studying BDNF in vitro offers valuable mechanistic insights but requires careful experimental design and rigorous controls due to:
Addressing these challenges often involves using complementary methods (e.g., combining ELISA with Western blot or ICC), validating reagents, and optimizing protocols tailored to the biological system being used.[11]
Table 4: Technical Challenges for BDNF.
|
Category |
Technical Challenge |
Explanation / Impact |
|
Low Abundance & Secretion |
BDNF is produced in small quantities |
BDNF is secreted at picogram-to-nanogram levels in vitro, requiring highly sensitive detection methods (e.g., ELISA, MS) and often sample concentration. |
|
Isoform Complexity |
Presence of proBDNF and mature BDNF |
BDNF exists in precursor (proBDNF) and mature forms with opposing biological effects. Many assays do not distinguish between them, leading to data misinterpretation. |
|
Antibody Specificity |
Limited or variable antibody performance |
Some antibodies cross-react with related proteins or fail to differentiate between BDNF isoforms, impacting Western blot, ELISA, and ICC accuracy. |
|
Protein Degradation |
BDNF is highly susceptible to proteolysis |
Without proper protease inhibitors, BDNF may degrade during sample collection or processing, leading to underestimation of true levels. |
|
Sample Adsorption |
Loss of BDNF to plastic surfaces |
BDNF can adhere to culture dishes, pipette tips, and tubes, reducing measurable concentrations, especially in low-volume or serum-free media. |
|
Media Interference |
Components of culture media affect assay performance |
Phenol red, serum proteins, and antibiotics can interfere with colorimetric or fluorometric detection in ELISA and ICC. |
|
Translational Disconnect |
mRNA levels do not reflect protein output |
qRT-PCR may show increased BDNF transcripts, but protein may not be translated or secreted due to post-transcriptional regulation. |
|
Standardization Issues |
Lack of universal standards or controls |
Different kits, reagents, and protocols lead to inter-laboratory variability and make it difficult to compare results across studies. |
|
Cell Line Limitations |
Cell-type variability in BDNF expression |
Some cell lines naturally express low or no BDNF, requiring transfection or recombinant supplementation, which may not mimic physiological regulation. |
|
Assay Sensitivity vs Specificity |
Trade-off between detecting low levels and ensuring specificity |
Highly sensitive methods (e.g., ELISA) may detect trace levels but lack specificity; highly specific methods (e.g., MS) may miss low-abundance protein. |
|
In Vitro vs In Vivo Relevance |
Limited physiological context in vitro |
In vitro models lack systemic interactions and extracellular matrix components found in vivo, affecting BDNF signalling and secretion dynamics. |
|
Reproducibility and Batch Effects |
Variability in reagents, culture conditions, and assay kits |
Inconsistent results due to differences in antibody lots, cell passage number, or incubation conditions are common across many BDNF studies. |
|
Quantification of Secreted BDNF |
BDNF rapidly degrades or internalizes in culture media after secretion |
Secreted BDNF is short-lived and may be rapidly reabsorbed by cells, making it difficult to detect or measure without optimized collection timing. |
CONCLUSION:
Brain-Derived Neurotrophic Factor (BDNF) is a key neurotrophin involved in numerous critical processes in the central nervous system, including neuronal survival, synaptic plasticity, neurogenesis, and cognitive function. Its complex synthesis, maturation, and activity-dependent release highlight its tightly regulated role in neural development and adaptation. As a central player in both physiological and pathological conditions from learning and memory to neurodegenerative and psychiatric disorders BDNF serves as an essential biomarker and a potential therapeutic target.
In vitro systems have become indispensable for studying BDNF biology, allowing researchers to dissect its molecular signalling, assess pharmacological effects, and develop targeted interventions in a controlled environment. However, quantifying BDNF in vitro presents significant challenges due to its low abundance, presence in multiple isoforms (proBDNF and mature BDNF), and susceptibility to degradation. Analytical methods such as ELISA, Western blotting, qPCR, and immunocytochemistry each offer unique strengths but also face limitations in sensitivity, specificity, and reproducibility. Methodological differences in sample handling, antibody selection, and detection thresholds further complicate direct comparison of results across studies.
Overcoming these technical barriers requires standardized protocols, improved detection reagents, and the integration of more sensitive platforms such as mass spectrometry or single-molecule assays. Future advancements in in vitro modelling including 3D cultures, organoids, and microfluidic systems may further refine our understanding of BDNF's role in neural health and disease.
In conclusion, while significant progress has been made in the study of BDNF, continued efforts are needed to enhance the accuracy and reliability of its quantification in vitro. These improvements will be crucial for translating basic research into effective diagnostics and therapeutics targeting BDNF-related pathways in neurological and psychiatric disorders.
REFERENCES




Dasari Cherishma Sri, Bandi Aasritha, Galla Naga Deepika, Dr. Adusumalli Surendra, Quantification of Brain-Derived Neurotrophic Factor (BDNF) in In Vitro Systems: Methodologies and Technical Challenges, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 9, 922-946. https://doi.org/10.5281/zenodo.17078030
 10.5281/zenodo.17078030
10.5281/zenodo.17078030
