Department of Pharmacy, Global group of Institutes, Amritsar -143501, Punjab, India.
Among the hundreds of billions of beneficial microorganism that exist on and in humans, the vast majority are permanently deposited in the digestive tract shortly after birth. The "gut-brain axis" results from the microbiota's complex and specific interactions with the host, particularly the central nervous system (CNS). The secretion of many metabolites, such as signalling molecules, bacterial structural components, and short-chain fatty acids (SCFAs), enables communication among the stomach, intestinal microbiota, and the brain. The microbiome directly influences an individual's character, mood, and sleep patterns, operating in conjunction with the brain. The human microbiota is a varied assemblage of microorganisms that predominantly resides in the gastrointestinal system. Modulation of immune system is one of their most vital functions with the age progression. The gastrointestinal tract attains this segregation through a mix of immune cells, mucus, immunoglobulin A (IgA), antimicrobial peptides, and epithelial cells. This location harbours the greatest quantity of commensals. They are referred to as the "mucosal firewall" when functioning in conjunction with the immune system. This review demonstrates the role of bacteria in neurodegenerative and autoimmune illnesses by investigating the various mechanisms facilitating bidirectional communication between intestinal microbiota and the brain's mucosal immune defences. Various neurodevelopmental and neurodegenerative disorders, such as Autism spectrum disorder, multiple sclerosis, Amyotrophic lateral sclerosis, anxiety, stress, and Parkinson's disease, are significantly affected by changes in microbiota composition.
Since 2007, genomes of 500 unique bacterial species have been identified, constituting the microbiome or microbiota that resides in the human intestine. The human colon is home to a vast and diverse microbiotaand the overall quantity of microorganisms in the human digestive system surpasses 100 billion [1]. Bacteria(Escherichia coli), viruses (Norovirus), fungi (Candida albicans), and archaea (Methanobrevibacter smithii) constitute the "gut microbiota" present in the human digestive system. The concept has been proposed that individuals are "meta-organisms" with bacterial populations that exceed their own cells by a factor of 10 to 100[2]. An individual's microbiota can be affected by age, sex, race, and several factors such as nutrition, drugs (particularly antibiotics), stress, smoking, gastrointestinal problems, and genetic predispositions[3]. Fermentation of undigested carbs, synthesis of short-chain fatty acids (SCFA), production of vitamins B and K, metabolism of medications, bile acids, and sterols, and provision of defence against external diseases are all ways in which different populations of microbes contribute to human health[4].The process of human gut bacterial production is inherently unique. The human fetus's stomach is devoid of germs or pathogens at the time of birth. During a child's development, several microorganisms inhabit the body. A multitude of factors affects the composition of a woman's microbiota, including the age of childbirth, the delivery method, dietary choices, hygiene practices, and antibiotic exposure[5]. Any interruption in the composition of the gut microbiota, known as dysbiosis, has been associated with an increased risk of neurodegenerative, immunological, neuropsychiatric, and metabolic diseases. The individuals who are genetically susceptible to certain diseases including amyotrophic lateral sclerosis, multiple sclerosis, and Parkinson's disease might develop dysbiosis as a result of the presence or absence of particular beneficial bacteria in their gut microbiota.
Regarding the qualitative composition, there is a growing body of literature attempting to associate distinct bacterial species with various physiological conditions. Microorganisms include Eubacterium, Faecalibacterium, Bacteroides, Bifidobacterium, Clostridium groups XIVa and IVa (which produce butyrate), Lactobacillus, and Roseburiaare thought to enhance the gut-brain axis, inflammation, immunology, metabolic condition, and cognitive performance[6]. Recent research has redefined the human-microbiota interaction as mutualistic, wherein bacteria assist our bodies with metabolic processes and immune system homeostasis that are not encoded in human genome. This study regards the microbiota as a crucial virtual organ in relation to pathophysiology and immune responses. Gut microbiota's role in neurological disorders and infectious diseases has been the subject of substantial research.
The gut microbiota contributes to immune system maintenance, fibre degradation, and carbohydrate metabolism. Moreover, gut microbiota have been linked to the regulation of neurophysiologically governed behaviours, including pain, multiple sclerosis, stress, and autism. The gut microbiota can modulate neurophysiological behaviours by affecting immunological, endocrine, and neurological pathways. It is now commonly known that the gut and brain work in tandem[7]. The gut microbiota composition is influenced by behavioural modifications as well environmental and host-related factors affect the microbial community[8]. The composition of gut microbiota correlates with modifications in behaviour and cognition, hence maintaining a balanced microbiome is critical for controlling the microbiota-gut-brain axis[9].
The afferent and efferent autonomic channels allow communication between the central nervous system and the gut's smooth muscle and mucosal layer, facilitating the gut-brain axis, the relationship between the central nervous system and the digestive tract. In response to environmental, such as emotional or stressful situations, the hypothalamus triggers the release of corticotrophin releasing hormone, which in turn activates the HPA axis. Gut bacteria produce neuroactive substances and metabolites that may influence the pathophysiology of some neurodegenerative diseases these include MS, ALS, Parkinson's disease, and Alzheimer's disease[10].
Bacteria present in the human gastrointestinal system are categorised based on their genus, family, order, and phylum. The intestinal microbiota, comprising the collective genetic material of all gut bacteria, predominantly consists of the phyla Bacteroidetes and Firmicutes, which together represent 90% to 95% of the entire microbial population[11]. Among the approximately 200 species that make up the phylum Firmicutes are Clostridium (representing 95% of the phylum), Blautia, Faecalibacterium, Enterococcus, Roseburium, Ruminococcus, and Lactobacillus Eubacterium.Prevotella and Bacteroides are the two most common genera in the Bacteroidetes phylum[12]. The members of the phyla Proteobacteria, Actinobacteria, Fusobacteria, Spirochaetes, and Verrucomicrobia are also prevalent in the gut microbiota, when these groups are out of equilibrium, dysbiosis emerges, which in turn causes metabolic abnormalities, irritable bowel syndrome, colon cancer, Crohn's disease, ulcerative colitis, and gastrointestinal problems[13]. Use cutting-edge methods that include DNA extraction and sequencing of the 18S, ITS, and 16S rRNA genes to determine and quantify the gut microbiota components. Although genetics offers extensive data on microbiome taxonomy, it yields minimal insight into the microbiome's essential activities[14].
Numerous variables influence the dynamic composition of gut microbiota. Factors encompass the anatomical position of the gut, mode of birth, age, weaning time, gestational age, antibiotic usage, ethnicity, food choices, and cultural influences. Helicobacter, Lactobacilli, and Veillonella are predominately occupied in the stomach, while Bacilli, Streptococcaceae, Actinomycinaeae, and Corynebacteriaceae are more common in the small intestine (duodenum, jejunum, and ileum). The quantity of Bacteroidetes and Lachnospiraceae in the colon escalates as one progresses up the ileum. Felicitative anaerobic microbes like Streptococcus, Staphylococcus, Bacteroidesfragilis, and Escherichia coli are part of the microbiota of babies born vaginally, along with Bifidobacterium longum and Bifidobacterium catenulatum. Similar to the microbiota of the mother's skin, infants born via caesarean section have fewer beneficial Bifidobacterium longum subspecies infantis bacteria.
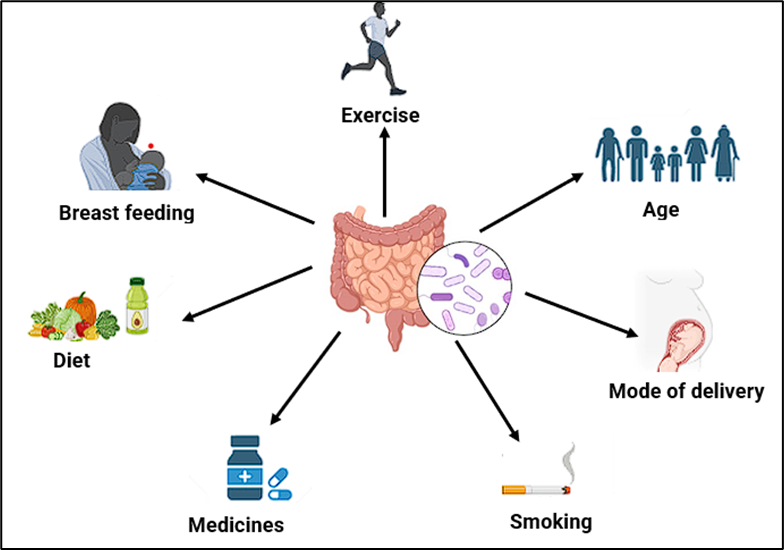
Figure 1 (a):Factors causing alteration in gut microbiota
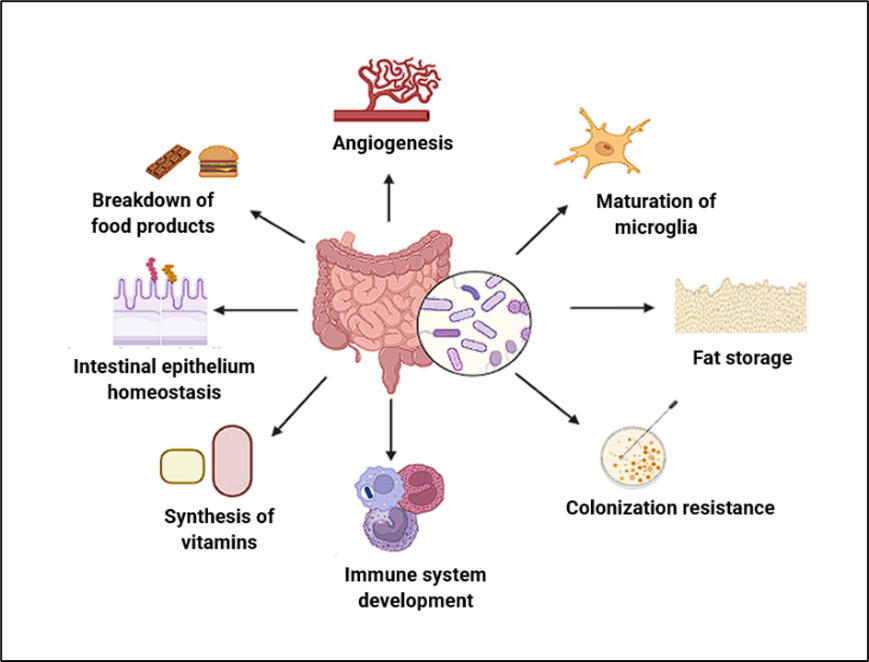
Figure 1 (b): Functions of gut microbiota
The host immune system isgreatly impacted by the microbiota in terms of its development, activation, and functionality. In a healthy individual, the symbiotic interaction between the gut mucosa and the microbiota is essential for immunological, metabolic, and gut-protective functions[15]. The gut microbiota functions as a distinct organ, exhibiting extensive metabolic capacity and significant functional adaptability.
1.3.1 Nutrient metabolism
Carbohydrates are essential for sustaining a healthy gut microbiome. Colonic microbes such as Bacteroides, Roseburia, Bifidobacterium, Fecalibacterium, and Enterobacteria produce butyrate, propionate, and acetate, among other short-chain fatty acids (SCFA), by fermenting indigestible oligosaccharides and carbohydrates that evaded proximal digestion[16]. These SCFAs have the potential to provide the host with a lot of energy. It is believed that the G protein-coupled receptor Gpr41 regulates the host energy balance through interactions with short-chain fatty acids. Peptide Tyrosine Tyrosine/Pancreatic Peptide YY3-36 (abbreviated as PYY) is another enteroendocrine hormone tied to this role[17]. Enzymes like polysaccharide lyases, glycoside hydrolases, and glycosyl transferases enable the majority of Bacteroides species to engage in carbohydrate metabolism.
1.3.2 Drug metabolism
Over 40 years ago, it was discovered that the gut microbiota can metabolise xenobiotics and pharmaceuticals. Studies indicate that p-cresol, a metabolite generated by gut bacteria, can impede hepatic sulfotransferases, hence complicating the function of liver to metabolise paracetamol. Recent studies indicate that digoxin and other cardiac glycosides up-regulate an operon including cytochrome, resulting in the inactivity of the ubiquitous Actinobacteria phylum bacterium Eggerthella lenta[18].
1.3.3 Protection from microbes
The gut mucosal immune system is subjected to significant stress in its endeavour to maintain homeostasis, which is contingent upon a well-balanced gut microbiota. Its principal role is to restrict the dissemination of endemic illnesses while concurrently accommodating advantageous commensals[19]. The two-layer mucus barrier, mostly formed in the large intestine, serves as a crucial antimicrobial defence mechanism that inhibits luminal microorganisms from contacting epithelial cells.
Furthermore, the gut microbiota has developed mechanisms to restrict the development of pathogenic bacteria by enhancing localized immunoglobulin synthesis. Previous studies indicate that some gut bacteria, particularly gram-negative species such as Bacteroides, may stimulate the release of secretory IgA by intestinal dendritic cells (DCs)[20]. The sIgA may subsequently cover the intestinal flora. The predominant portion of sIgA that encompasses the microbiota belongs to the sIgA2 subclass, as it exhibits greater resistance to degradation by bacterial proteases[21]. In addition, intestinal epithelial cells (IECs) can produce a bacterial-detection ligand through a Toll-like receptor mediated pathway; this ligand then helps to switch from systemic sIgA1 to intestinal mucosal sIgA2.These systems keep bacteria from migrating from the intestinal lumen to the bloodstream, which limits an immune response on a systemic level[22].
The microbiota and both the innate and adaptive immune systems are all part of gut immunomodulation. Many different kinds of immune system cells and components are involved in immunomodulatory activities in the gut [23]. These include GALT, effector and regulatory T cells, IgA-secreting B (plasma) cells, resident macrophages, Group 3 innate lymphoid cells, and dendritic cells in the lamina propria. Impairments in the development of Peyer's patches and isolated lymphoid follicles, as well as the prevalence of IgE+ B cells over the normally seen IgA+ B cells, point to the importance of the gut microbiota in the formation of functional gut-associated lymphoid tissue (GALT). In the gut, effector T cell responses are regulated by Th2 responses instead of Th1 responses, according to the research. In a normal setting, Th1 and Th17 cells mostly control Th1 responses[24]. Innate lymphoid cells (ILCs) are a subset of innate immune cells that produce immediate response to signals fromthe epithelium in the form of cytokines. In contrast to other types of T helper cells, ILCs originate from lymphoid progenitors and develop into cytokine-producing cells, specifically Th17 cells[25]. It appears that RORγt+ ILCs are more intimately linked to the modulation of intestinal immunity [26]. An effective collaboration between the immune system and microbiota selectively modulates and concludes reactions reciprocally, coordinating the body's innate and adaptive immune responses. The development of a complex immune system, as well as the reliance on bacteria, came with certain drawbacks.The absence of regulation over aberrant immune reactions to self, microbiota, or environmental antigens is the fundamental cause of various human illnesses, including inflammatory disorders, allergies, and autoimmune diseases. Antibiotic usage, dietary changes, and the recent eradication of natural partners like helminth worms have altered the composition and function of the microbiota, which has the potential to deteriorate the gut microbiome. Microbiota, commonly referred to as commensals, engage in various symbiotic relationships with their mammalian hosts, characterized by continuous interactions[27].
The immune system normally encounters commensals for the first time during birth, since the foetal gastrointestinal tract is believed to be sterile. It is clear that components in breast milk may impact some of these initial responses to commensals; nevertheless, the understanding of how newborn tissues adjust to the huge challenge of microbial colonisation is still inadequate[28]. Colostrum and breast milk include a variety of components, including cytokines, immune cells, metabolites, immunoglobulin A, and live microorganisms. Maternal IgA binds to nutritional and microbial antigens, limiting immunological activation and microbial adherence; metabolites, especially oligosaccharides, in breast milk encourage the growth of some bacteria, like Bifidobacterium. Dendritic cells in milk, which are enriched with bacteria, may modulate the immune response to commensal antigens, potentially resulting in newborn immunological imprinting[29].
Perhaps the most important function of the human microbiome is the microbiota's involvement in immune system development and maturation. The process of microbial colonisation of the host commences throughout gestation and accelerates significantly once delivery[30]. The microbiota in humans instructs the immune system to manage diverse commensal antigens, sustains immunological homeostasis, and inhibits dysregulated inflammation. It instructs intestinal epithelial cells and the associated immune cells on how to appropriately respond to microbial signals[31]. A category of human disorders termed "the hygiene hypothesis" posits that insufficient exposure to pathogenic agents and beneficial microbes, such as gut microbiota, markedly increases the incidence of autoimmune and allergy diseases, particularly in infants. Significant epidemiological statistics globally support this theory. Consider Finland, a highly advanced nation, as an illustration. Type 1 diabetes mellitus (T1DM) is six times more common in that area than in the less developed neighbouring Karelian Republic of Russia. Type 1 diabetes mellitus (T1DM) is an autoimmune disorder that causes the destruction of the pancreatic β-cells resulting in the abnormal blood glucose levels. A plausible reason for the significant gap in type 1 diabetes prevalence is that children in underdeveloped areas are more frequently exposed to a wider array of microbial antigens at an earlier age. Exposure to hookworms and other endotoxins during the first year of life may confer protection against the development of asthma, a disorder more prevalent in urban environments. Thus, a stable and healthy gut microbiota, together with early exposure to a variety of benign bacteria, is essential for an individual's overall well-being[32]. An important part of the immune system's inherent function is the regulation of microbiota. Skin and the gastrointestinal tract, which are both home to commensals, therefore have the body's largest concentration of immune cells. The gastrointestinal tract attains this segregation through a mix of immune cells, mucus, immunoglobulin A (IgA), antimicrobial peptides, and epithelial cells. They are referred to as the "mucosal firewall" when functioning in conjunction with the immune system[33].
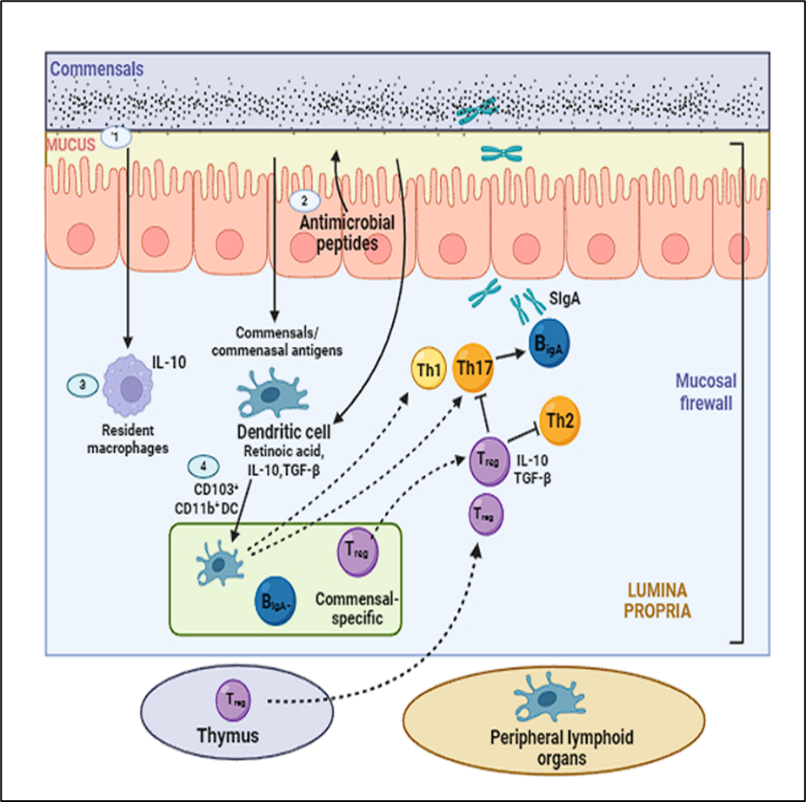
Figure 2: Mucosal Firewall
(1) The primary barrier preventing germs from migrating to host tissues is mucus, which restricts their contact.(2)An additional crucial element in minimizing interaction with commensal bacteria is the antimicrobial peptides synthesized by epithelial cells. (3) The indigenous macrophages in the tissues swiftly destroy the translocating commensals. (4) Although they do not travel further, CD103+ CD11b+ dendritic cells can acquire commensals or commensal antigens en route to the mesenteric lymph nodes from the lamina propria.Th17 cells, B cells that secrete IgA, and Tregs, which are specific to commensal antigens, differentiate after these dendritic cells present them.The "mucosal firewall" is a complex system of cells and tissues that protects the stomach's lymphoid tissues from commensals by limiting their access to these tissues. This system includes the epithelium barrier, the mucus layer, IgA, dendritic cells, and T cells[34].
3.1 Role of metabolites produced by the gut microbiota in the brain
A diverse array of metabolites is generated by numerous bacterial genera and species in conjunction with their hosts. The connections between intestinal and central nervous system are facilitated by a variety of bacterial compounds[35]. Gut bacteria may mediate communication between the central nervous system and the brain through the release of certain active metabolites. Microflora components such as bile acids, aromatic amino acids, and short-chain fatty acids (SCFAs) mostly impact the brain. Bacterial fermentation of carbohydrates generates propionate, butyrate, and acetate, which are the primary constituents of saturated fatty acids[36]. Acetate is the most abundant short-chain fatty acid in the blood, which helps maintain a healthy colon because the liver mostly breaks down propionate and the epithelial mucosa handles butyrate metabolism. These acids might improve their interactions with the gut by binding to receptors that are connected to G-proteins. When SCFAs enter cells, they bind to G protein-coupled receptors GPR41, GPR43, and GPR109A, which are found on immune cells and gut epithelium, respectively[37]. Another strategy is to intracellularly suppress nuclear class I histone deacetylases (HDACs), namely HDAC1 and HDAC3. Impediment of HDACs is linked to most anti-inflammatory immune features, such as reduced levels of proinflammatory cytokines (e.g., IL-6, IL-8, TNF-α) and NF-κB activity

Figure 3: Regulation of gut metabolites in the brain
Abbreviations: MUC2- Mucin 2, SCFA- Short chain fatty acid, HDAc- Histone acetylation, EEC- Enteroendocrine cell, GLP-1 Glucagon-like Peptide 1, PYY- Peptide YY, pAMPK- Phosphorylated Adenosine monophosphate activated protein kinase, AMP- Adenosine monophosphate, ATP-Adenosine triphosphate, LEPR- Leptin receptor.
The microbiome of the large intestine metabolizes indigestible carbohydrates and dietary fiber to generate short-chain fatty acids, namely butyrate, propionate, and acetate.Anti-inflammatory medicines hinder the maturation of naive T cells into regulatory T cells by blocking HDAC. This leads to a decrease in the production of cytokines that promote inflammation, such as IL-6, IL-8, TNF-α, and NF-κB activity. Short-chain fatty acids boost intestinal gluconeogenesis thus causing glucose tolerance. Additionally, Enteroendocrine cells can be stimulated to release the hunger-inhibiting hormones GLP-1(Glucagon-like Peptide1) and PYY by short-chain fatty acids (SCFA).Adipose tissue produces increased levels of leptin in response to exposure to short-chain fatty acids, resulting in diminished appetite.
3.2 Gut microbiota and their impact on the central nervous system: Direct and indirect effects
Potentially affecting levels in the brain are the precursors of neurotransmitters that are produced in the stomach. Some neurotransmitters may originate in the digestive process, but bacteria can also synthesise them[38]. Microorganisms can influence host brain function in various ways; for instance, Escherichia coli can emit dopamine, serotonin, and noradrenaline, and Lactobacilli can produce serotonin, GABA, acetylcholine, and histamine[39]. This process has been associated with a wide range of neurological illnesses, such as depression, autism spectrum disorders, Alzheimer's disease, and Parkinson's disease. Additionally, enteroendocrine cells can stimulated to secrete specific gut hormones, such as leptin and glucagon-like peptide-1 (GLP-1), by short-chain fatty acids (SCFAs), which could have an indirect effect on the gut-brain axis[40]. The vagus and enteric nerve networks constitute components of the neurological system, whereas cytokines and cortisol are integral to the immune system. The brain utilises analogous mechanisms to modify the gut microbiome composition in reaction to stress. The hypothalamus-pituitary-adrenal axis regulates cortisol release, which affects immune cell secretions both locally and systemically, including cytokine production from the intestines. Furthermore, cortisol possesses the ability to alter the composition of gut bacteria, therefore affecting gut permeability and barrier function. The transmission of effects from the gut microbiota to the brain is intricately related to the vagus nerve and the regulation of systemic tryptophan levels.

Figure 4: Communication between gut microbiota and brain
3.3 Microbiota role in the brain development
There are a number of neurological processes that the gut microbiota can influence, including microglial activity and capacity, myelination, neurogenesis, blood-brain barrier formation and stability, and more. The utilisation of germ-free mice has enabled researchers to elucidate the significance of the microbiome in the initial phases of brain development[41]. Using a genome-wide transcriptomic approach, researchers found that germ-free mice showed elevated gene expression in areas responsible for a variety of metabolic and adaptive functions, such as steroid hormone metabolism, cyclic adenosine 5-phosphate-mediated signalling, and long-term synaptic potentiation; the area’s most impacted were the hippocampus and cerebellum[42]. A combination of probiotics and exercise can restore neurogenesis that has been diminished by antibiotic treatment; nevertheless, in germ-free mice, adult dorsal hippocampus neurogenesis is boosted. Changes in locomotor and exploratory behaviour are brought about by increased turnover of dopaminergic and serotonergic neurotransmitters and heightened synaptogenesis markers in the striatum of germ-free mice[43]. The basolateral amygdala mediates anxiety, fear, and social behaviour, with germ-free mice displaying larger volumes than their germ-infested counterparts. The research also identified dendritic hypertrophy in this region. Mice lacking microorganisms exhibited hypermyelination and increased expression of myelination-associated genes in the prefrontal brain[44]. Comparing the microbiome of individuals with high and low growth phenotypes revealed that those with high growth exhibited reduced levels of neurofilament-L, MBP, and NeuN, which are all indicators of myelination[45].
Recent data suggests that several neurodegenerative illnesses may involve neuroinflammation as a pathogenic factor. Proinflammatory substances like TNF-α, IL-6, or MCP-1 are released by activated microglia in a neuroinflammatory condition, combined with reactive oxygen species made by glial cells and resident macrophages, leading to chronic neuroinflammation. It is becoming more and more apparent that changes to the naturally occurring microbiome could have a role in the onset of neurodegenerative illnesses, some neuropsychiatric disorders, and other neurological conditions[46]. Neurodegeneration can be linked to a persistent reduction in microbial diversity. Modifications to the gut microbiota are mostly linked to chronic inflammation and the deterioration of intestinal epithelial cell integrity; nevertheless, they may also induce neuroinflammation, neurodegenerative disorders, and neuropsychiatric conditions. The microorganisms impede brain cell death and diminish oxidative stress by downregulating the TLR4/NF-κB signalling pathway. Fig 5 illustrates the association between gastrointestinal abnormalities and central nervous system issues; later sections elaborate on this subject[47].

Figure 5: Role of dysbiosis in various neurodegenerative disorders
3.5 Parkinson's disease
Among the most common neurodegenerative diseases, Parkinson's may have its roots in imbalances in the microbiota, which in turn affect the brain. The progressive and long-lasting disorder known as Parkinson's disease is characterised by both motor and non-motor symptoms[48]. The prevalence rises with age and usually affects those above the age of 40, irrespective of gender. The multifocal neurodegenerative condition known as Parkinson's disease (PD) is caused by the buildup of alpha-synuclein (α-syn) in the neuronal cell body, which is called progressive alpha-synucleinopathy[49]. This eventually results in the formation of Lewy bodies, which are spherical, eosinophilic cytoplasmic inclusions exhibiting lamellation. Tremors and bradykinesia are the most common motor symptoms, while cognitive deterioration, sleep problems, depression, and anxiety are the most common non-motor symptoms[50]. Some examples of conditions that do not involve movement include dementia, depression, impaired smell, and problems with the digestive system including nausea, difficulty swallowing, excessive salivation, constipation, and problems with defecation.
The aetiology of Parkinson's disease may be influenced by an imbalance in the gut microbiota, which can lead to changes in intestinal motility, increased permeability, and chronic local inflammation[51]. Researchers have looked at the gut microbiome composition of individuals suffering Parkinson's disease considering gastrointestinal dysfunction is a defining feature of the disease. Individuals with Parkinson's disease often have an overabundance of the Enterobacteriaceae bacteria, which has been linked to postural instability in studies.
The intensity of postural instability and gait impairment has been significantly associated with the relative abundance of Enterobacteriaceae. The cell wall of gram-negative bacteria generates serum lipopolysaccharide (LPS), which escalates with the proliferation of Enterobacteriaceae bacteria. The intestinal epithelium barrier is compromised when neurotoxins such as LPS infiltrate the bloodstream via the gut wall[52]. Systemic inflammation can be induced when lipopolysaccharides (LPS) in the bloodstream produce inflammatory cytokines through the toll-like receptor (TLR) 4 and nuclear factor-kappa B (NF-κB) pathway. The disruption of the blood-brain barrier and the facilitation of α-synuclein deposition are caused by LPS and inflammatory cytokines, such as tumour necrosis factor (TNF-α), interleukin (IL)-1β, and IL-6. According to FAO/WHO, "Live microorganisms that, when administered or ingested in sufficient quantities, provide a health benefit to the host." Probiotics, which include bifidobacteria and lactobacilli, have shown promise in reversing conditions that are similar to Parkinson's disease.
3.6 Alzheimer's disease
The most common neurological condition is Alzheimer's disease. Degenerative brain changes caused by Alzheimer's disease (AD) don't manifest themselves for decades, hence the disease disproportionately affects the elderly[53]. Aβ plaques, created by APP cleavage, and neurofibrillary tangles (NFTs), made of hyperphosphorylated tau protein, are disease markers that accumulate in the brain[54].
Soluble oligomers of Aβ and/or Tau protein are the main detrimental elements impacting brain tissue, as per an alternate theory. When the immune system erroneously perceives insoluble Aβ deposits as exogenous entities, it may trigger an inflammatory cascade, potentially resulting in neuronal damage. Microglia and astrocyte activation is the principal pathway by which neurofibrillary tangles and amyloid plaques cause brain inflammation[55].
During gut dysbiosis, which is linked to inflammation and increased intestinal permeability, microorganisms such as spirochaete, Herpes simplex virus type 1, Chlamydia pneumoniae, α-N-methylamino-L-alanine (BMAA), LPS, and microbial amyloids are found in the bloodstream at elevated levels. Intestinal cyanobacteria produce the neurotoxic BMAA, which causes neurodegeneration, cognitive impairments, astrogliosis, and neurofibrillary tangle formation in adult rats after exposure to the compound during neonatal period[56]. The production of Aβ fibrils, which are involved in neuroinflammation in the brains of Alzheimer's disease patients and a strong activator of NF-κB, is initiated by LPS. Microglia have the toll-like receptor 4 (TLR4) that interacts with lipopolysaccharide (LPS). Essential for TLR4 activity is the coreceptor CD14[57]. Microglia in Alzheimer's disease brains show increased levels of CD14, demonstrating the role of CD14 and TLR4 in the pathogenesis of Alzheimer's disease. The innate immune system could be activated by the amyloid produced by the gut microbiota, leading to a worsening of inflammation caused by cerebral Aβ42[58].It culminates in a chronic condition characterized by low-grade inflammation known as inflammaging[59]. Certain investigations suggests that the transfer of normal microbiota from control mice to Alzheimer's disease models in mice have further substantiated the existence of the gut-brain axis in the pathology[60]. The normalisation of gut flora resulted in a reduction of Aβ plaque and neurofibrillary tangle generation, a decrease in glial reactivity, and an enhancement of cognitive function[61]. Pathological changes in microbiota composition, such Clostridium difficile overgrowth and reduced Bifidobacteria, can alter the expression of proinflammatory molecules[62].
3.7. Multiple sclerosis
A chronic immune-mediated disease of the nervous system, multiple sclerosis (MS) manifests over time. Multiple sclerosis is defined by neurodegeneration, demyelination, and axonal damage. Symptoms that may be experienced include vertigo, blurred vision, lightheadedness, pain, diminished motor skills, lack of coordination, fatigue, tingling, depression, and cognitive deterioration[63]. The majority of people who suffer from multiple sclerosis (MS) are women, and the disease mainly affects younger persons, in contrast to Parkinson's disease and Alzheimer's disease. It affects the central nervous system and is characterised by inflammation and demyelination. Relapsing-Remitting MS (RRMS), Progressive-Relapsing MS (PRMS), Primary Progressive MS (PPMS), andSecondary Progressive MS (SPMS) are the main types of multiple sclerosis[64]. Several factors, both genetic and environmental, impact the pathophysiology of multiple sclerosis. The predominant and thoroughly proven causes encompass childhood obesity, insufficient blood vitamin D levels, inadequate sun exposure, and smoking. All of these variables can also indirectly influence the gut microbiome[65]. The gut microbiota plays an important role in the development of MS because certain bacteria have the ability to affect T-cells and, by extension, the immune system[66]. The etiology of multiple sclerosis remains enigmatic, however a weakened immune system is commonly regarded as a significant factor[67, 68]. Multiple sclerosis not only induces chronic inflammation but also results in a compromised blood-brain barrier that exhibits reduced selectivity in the brains of affected individuals[69]. The animal model of demyelinating illnesses, experimental autoimmune encephalomyelitis (EAE), was worsened by the transplantation of microbiota from multiple sclerosis (MS) patients to germ-free mice, in contrast to germ-free animals given healthy microbiota[70].This toxin induces autoimmune demyelinating phenomena and injures neurones and oligodendrocytes by compromising the blood-brain barrier[71]. Species of Dorea capable of degrading mucin, metabolising sialic acids, and activating IFNγ are quite uncommon. Dorea enhances the proliferation of Blautia by generating gases that the latter can utilise. Reports indicate a heightened prevalence of Akkermansia in multiple sclerosis patients[72]. Blautia may enhance cytokine production that fosters inflammation and degrade mucin[73]. Another study revealed that levels of Adlercreutziaequolifaciens remained unchanged in MS patients, although levels of Collinsella and Slackia diminished. Sutterella (Proteobacteria) was present in reduced abundance in multiple sclerosis, but Mycoplana, Pseudomonas, Bilophila, and Acinetobacter calcoaceticus exhibited increased prevalence. Inflammatory cytokines generated by Acinetobacter calcoaceticus and a reduction in regulatory CD4 T cells can aggravate multiple sclerosis[74].
3.8. Amyotrophic lateral sclerosis
Degeneration of motor neurones, spinal cord, and brain is a hallmark of amyotrophic lateral sclerosis (ALS), a neurological disease. Symptoms may present as diminished strength, cramps, impaired coordination, rigidity, spasms, or muscle twitching. Difficulties in speaking, swallowing, and breathing manifest progressively as a consequence of this[75]. Three distinct phases characterise amyotrophic lateral sclerosis (ALS) in animal models: a) pre-onset (60-70 days), b) onset (90-100 days), and c) progressive (120-130 days)[76].During the pre-onset period, transgenic mice exhibit increased intestinal permeability and disruption of the blood-brain barrier. Intestinal wall damage is associated with a decrease in E-cadherin and zonula occludens (ZO)-1. There was a decrease in the quantity of butyrate-producing bacteria like Peptostreptococcus and Butyrivibrio fibrosolvens in the bloodstream, and the levels of the pro-inflammatory cytokine IL-17 were higher in the intestines and serum of SOD1G93A animals. According to Ruhé H G et.al the amyotrophic lateral sclerosis (ALS) was slowed down in SOD1 transgenic mice when 2% butyrate was added to their drinking water [77]. Several species of Dorea are enhanced in the faeces of ALS patients, but the numbers of butyrate-producing bacteria, such as Oscillibacter, Anaerostipes, and Lachnospira, are lowered. Evidence suggests that BMAA and LPS plasma levels are altered in amyotrophic lateral sclerosis (ALS) patients[78]. When these substances reach the bloodstream, they have the potential to cause neuroinflammation and weaken the blood-brain barrier, two factors that contribute to the development of amyotrophic lateral sclerosis (ALS)[79].
3.9. Major depressive disorder
Major depressive disorder (MDD) is a significant contributor to disability, morbidity, and mortality in industrialised countries. In 2017, Major depressive disorder affected around 264 million individuals worldwide. Distress, impaired concentration, profound fatigue, alterations in appetite, and difficulties with sleep and digestion are characteristic features of major depressive disorder [80]. A potential underlying cause of the condition is a deficiency of monoamine neurotransmitters such as serotonin, dopamine, and noradrenaline[81]. The concept that depression is a systemic illness is supported by the fact that high blood cytokine levels, which indicate systemic inflammation, are another possible cause of the illness. Contributing to the development of major depressive disorder and on behaviour and emotions, systemic inflammation produces neuroinflammation and activates microglia and astrocytes[82].Considering knowledge has been discovered about gut microbiota and chemicals derived from bacteria, researchers have pinpointed disruptions in the gut-brain axis as a distinct component of the pathophysiology of major depressive disorder[83]. Another study found that the gut microbiome of depressed patients differed significantly from that of healthy controls. A study involving animals has demonstrated analogous findings, revealing an elevation in Bacteroidetes and a reduction in Firmicutes across several depression models[84]. Furthermore, one more research indicates that patients self-assessments of their health, physical capabilities, energy, mental well-being, and social functioning exhibit a favourable correlation with the presence of the advantageous short-chain fatty acid butyrate, generated by the bacterium Coprococcus, in their gastrointestinal tract.
3.9. Autism spectrum disorder
Neurodevelopmental disorders categorised as "autism spectrum disorders"encompass restricted, repetitive, and stereotypical interests and behaviours, as well as challenges in social interaction and communication. It has been suggested that gastrointestinal problems, maybe linked to an atypical composition of the gut microbiota, may play a role in ASD, as gastrointestinal symptoms are frequently found in children with ASD[85]. The gut microbiota composition can be influenced by a number of factors, including dietary differences and increased antibiotic use in ASD patients compared to neurotypical controls; hence, care must be used when interpreting these results. A study has indicated alterations in the amounts of short-chain fatty acids in the stools of children with autism spectrum disorder [86]. This suggests that bacteria affect brain function by modifying the synthesis of their neuroactive metabolites. Individuals with ASD frequently encounter gastrointestinal complications, including diarrhoea, constipation, and abdominal discomfort, alongside the psychological dimensions of the condition. Several studies indicate that persons with ASD exhibit elevated levels of specific microorganisms, including Clostridium and Lactobacillus. A study indicated that the intestinal microbiota of autistic children exhibited reduced levels of the beneficial species Prevotella and Coprococcus in comparison to healthy controls. Moreover, adult autistic patients had significantly elevated blood concentrations of bacterial LPS and inflammatory biomarkers IL-1β and IL-6 in comparison to healthy controls. The pathophysiology of autism is insufficiently understood to make definitive assertions regarding the impact of low-grade endotoxemia[87].
CONCLUSION AND FUTURE PERSPECTIVE
The relationship between microbiota aberrations and pathological changes remains ambiguous; however, researchers have increasingly focused on gut microbiota due to its significance for human function. Nevertheless, the profound importance of a balanced gut microbiota composition and the synthesis of many bacterial metabolites for human health, encompassing the central nervous system and immune system, has been shown. The gut-brain axis is a complex web of relationships between the CNS, the metabolic processes of gut bacteria, and the enteric nervous system, all of which may play a role in the development of neurological diseases. In recent decades, there has been a noticeable rise in the occurrence of autoimmune illnesses, which are characterised by dysregulated immune responses andin immunology, a fascinating junction has formed. A complex and dynamic microbial community inhabits the human intestines. The gut microbiota greatly influences food digestion, immunopotentiation, microvilli formation stimulation, dietary fibre fermentation, and pathogen colonisation prevention. Numerous animal and observational research indicate that gut microbiota may alter gut-brain-axis function, hence influencing the neuropathogenesis of central nervous system disorders. Multiple sclerosis, Amyotrophic lateral sclerosis, Parkinson's disease, and Alzheimer's disease are among the neurodegenerative conditions that can be accelerated by increased levels of bacterial metabolites and inflammatory cytokines caused by dysbiosis can affect the permeability of gut and the blood-brain barrier, leading to neuroinflammation. Probiotics could provide protection against a range of neurodegenerative disorders by preventing intestinal dysbiosis.
CONFLICT OF INTEREST
The authors declare no conflict of interest.
REFERENCES

Sandip Tejpal, Mayank Attri, Akash Sharma, Ritika Puri, Manbir Kaur, Lavish Kumar, Tejinder Singh, Inderjeet Singh, The Gut-Brain-Immune Axis: Linking Microbiota to Autoimmunity and Neurodegeneration, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 12, 4178-4198. https://doi.org/10.5281/zenodo.18099010
 10.5281/zenodo.18099010
10.5281/zenodo.18099010
