
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
Sri Raghavendra College of Pharmacy, Chitradurga, Karnataka, India 577501.
Punicalagin it is an Ellagitannin compound that found naturally in Punica granatum or pomegranate, it possesses strong antioxidant effects through several biological pathways. It is known to neutralize a byproduct of mitochondrial metabolism reactive oxygen species (ROS), and suppresses the activity of NADPH oxidase, also binds with transition metals like Fe²? (ferrous) and Cu²? (copper), which helps block the production of harmful hydroxyl radicals. In vascular endothelial cells, punicalagin promotes the synthesis of nitric oxide (NO) by activating eNOS or endothelial nitric oxide synthase through a calcium dependent mechanism. Nitric oxide then plays a further protective role by decreasing ROS levels acting as a superoxide scavenger, maintaining eNOS in its functional form, and improving gene expression related to oxidative balance. These combined effects help to protect the cells against oxidative damage and maintain redox stability. This mechanistic understanding supports the potential of punicalagin as a t Medicinal substance in conditions due to oxidative stress.
Reactive oxygen species (ROS) are chemically reactive oxygen molecules can readily engage in various cellular reactions. that contain both unstable oxygen molecule and free radicals and produced during mitochondrial respiration [1,2]. These reactive species can affect the Overwhelming the body's natural antioxidant defenses, which results in oxidative stress and associated cellular injury [2]. In our body ROS can be formed by both non-enzymatic and enzymatic mechanisms [3]. Under physiological conditions, they are byproducts of normal metabolic processes, especially in reactions that take place in the mitochondrial electron transport chain. Apart from their effective role in biotransformation. Unstable oxygen molecules or Reactive oxygen species (ROS) are involved in essential cellular functions., including proliferation and differentiation of cells, both inner side and outer side of the cell [4]. However, when their elevated production affects capacity of antioxidant defenses; ROS can cause significant cellular injury or trigger protective pathways aimed at mitigating their effects [5]. Several compounds of pomegranate Compounds such as punicalagins and punicalins, have demonstrated about their effective antioxidant activity. Notably, punicalagin has been shown in multiple studies to possess effective free radical scavenging properties, largely due to its high hydroxyl group content [6].
Punicalagin
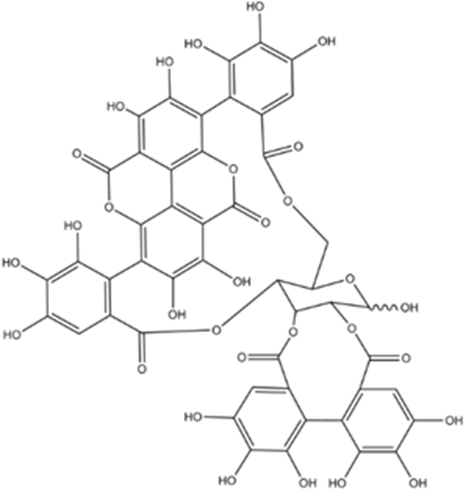
Figure 1. Chemical Structure of Punicalagin
Punicalagin is an important ellagitannin naturally occur in pomegranate [7]. It is a large, water miscible, hydrolysable tannin that exists in two structural forms alpha and beta isomers. This polyphenolic compound is not exclusive to pomegranate; it is also found in other plant species within the Myrtales order, including Indian almond. In pomegranate, punicalagin is mainly present in the peel, arils, husk, juice, and other parts of the fruit [8].
Source of punicalagin:
A polyphenolic compound punicalagin found commonly in different parts of the Punica granatum, especially in fruit and husk, as well as in the leaves of Lafoensia pacari a tree belongs to Lythraceae family. It has two structural isomers, alpha and beta. Additionally, punicalagin has been detected in several species within the Terminalia genus, including Terminalia. arborea, T. arjuna, leaves of T. brachystemma , bark of Terminalia. brownie, Roots of T. laxiflora, Terminalia. calamansanai, fruits of Terminalia. chebula, Terminalia. citrina (fruit), Terminalia. ivorensis (bark), bark of T. macroptera, leaves of Terminalia. muelleri, and leaves of Terminalia. oblongata. [9].
In addition, punicalagin of pomegranate Commonly present in a variety of plant-based foods, including berries. including raspberries, strawberries (Fragaria species), and some amount also found in walnuts [10].
The husk of pomegranate, which remains as a major by-product after squeezing juice, is rich in several ellagitannins. Include ellagic acid, ellagitannin hexosides, punicalin and hexahydroxydiphenoyl (HHDP) etc [11].
Various research Studies described that pomegranate peel is the major by-product generated during juice production compared to other parts of plant, the flowers of pomegranate also contain high concentration of polyphenolic compounds including pedunculagin, punicalagin (PUN), and other tannins like ellagic acid, which meant for their effective potential for various applications [12].

Figure 2. Ellagitannin compounds found in pomegranate
In addition to pomegranate, punicalagin is present in the leaves of Terminalia catappa, commonly referred to as tropical almond, which belongs to the Combretaceae family.
Traditionally, different parts of this plant—such as the shoot, leaves, fruit, and trunk have been utilized in Southeast Asian folk medical remedy. These parts have shown protective properties against genotoxic damage induced by substances like bleomycin [13].
Metabolism and bio availability of punicalagin:
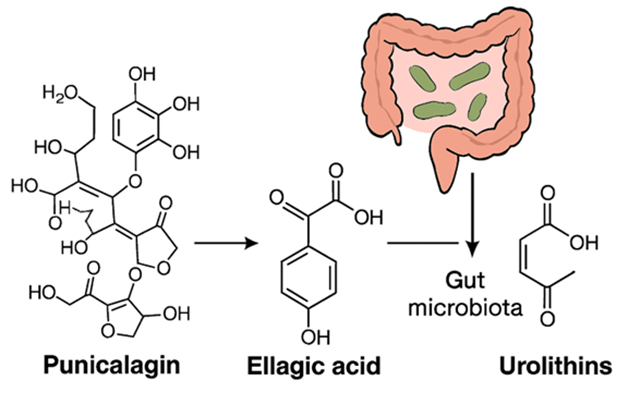
Figure 3. The metabolic processing of punicalagin (Detailed visual of punicalagin metabolism. It traces the production of tannin called as ellagic acid from punicalagin and then to urolithins, facilitated by gut microbiota.)
The biotransformation of ellagitannins in the GI tract involves multiple steps. Several Ellagitannins, such as punicalagin (PUN), are hydrolyzed in small intestine, releasing ellagic acid (EA) [14]. Despite this, both ellagitannins and EA have limited bioavailability. Those compounds that remain unabsorbed in the small intestine undergo further breakdown by gut microbiota [15]. Through a series of decarboxylation reactions, intestinal bacteria convert EA into several intermediate compounds, ultimately producing metabolites known as urolithins, which belong to the dibenzopyran-6-one structural class [16].
Due to this complex metabolic process, it is thought that urolithins, rather than punicalagin (PUN) or ellagic acid (EA), are the primary biologically active pounds. Ellagic acid is metabolized into urolithins, through the opening of one lactone ring followed by the removal of hydroxyl groups. Structurally, these metabolites combine features of coumarin and Iso coumarin and are also referred to as benzo coumarins [15].
The end products of this metabolic process including Iso-urolithin A, urolithin A&B. Interestingly, not all individuals can produce these end products, due to variability in gut microbial composition. Based on this, researchers have categorized individuals into three distinct urolithin metabolic profiles (UM): UM-A, where only urolithin A is produced; Urolithin A&B, UM-B, and Iso-urolithin A are generated in certain individuals, while others, classified as UM-0, do not generate any urolithins [17,18].Classification is made according to the urolithin profile observed in urine 24 hours following ingestion [16]. Human gut bacteria of the genus Gordonibacter are directly involved in the conversion of EA to intermediate urolithins [19,20]. A significant positive association was observed between these fecal bacteria and urolithin A concentrations in feces and urine. [21]. The abundance of Gordonibacter bacteria was greater in subjects classified as metabotype A than in those belonging to metabotype B or 0. [22] Numerous research has also explored genes encoding enzymes implicated in the metabolism of EA into urolithins, identifying potential candidate enzymes that may contribute to this process. [23]
The fast transformation of ellagitannins into ellagic acid within the gastrointestinal tract (GI) raises questions about the validity of using compounds such as punicalagin directly in in vitro culture studies [16]. The initial bioavailability studies of pomegranate ellagitannins in rats demonstrated that following high-dose ingestion, metabolites including urolithins A, B, and C, along with trace levels of ellagic acid dimethyl ether glucuronide, were present in plasma and urine [24]. Although trace amounts of punicalagin were observed in rats following long-term administration or ingestion, this result was not replicated in human trials. [25].
In research carried out by Gonzales and colleagues, healthy participants were given two types of pomegranate extracts that varied in their content of free ellagic acid (EA). The study also modeled gastrointestinal conditions, including variations in pH value and amount of protein to explore the factors influencing the Physiological accessibility of free ellagic acid. The study focused on how punicalagin is converted into EA, followed by the metabolism of EA into urolithins. The studies determined how changes in pH and protein presence influence the solubility and absorption of both Punicalagin and ellagic acid [18].
Numerous studies shown that EA reaches the highest concentration of approximately 100 nM in blood plasma within an hour after ingestion, even when taken in relatively larger doses. This level remains detectable in the bloodstream for about 5 to 24 hours following administration [15,18]. Additional experiments demonstrated that EA's solubility is significantly affected by pH, especially between pH 7 and 8, while its binding or interaction with proteins appeared to be minimal. On the other hand, PUN exhibited no notable change in solubility across different pH levels; however, in a protein-rich environment, it showed strong binding to proteins. Within this framework, PUN did not undergo hydrolysis to release EA, indicating that protein interactions may hinder its conversion [17,18].
Despite encouraging therapeutic results in preclinical models, both EA and PUN face limitations beacuse to their poor solubility, which hampers effective absorption after oral ingestion. Ellagic acid (EA) has been designated as a Class 4 compound based on the BCS (Biopharmaceutical Classification System) which means it has poor solubility and low intestinal permeability, limiting its clinical applications [26].
To enhance the bioavailability of EA and PUN, one approach involves reducing their particle size through micronization techniques [27]. Typical techniques for nanoparticle production involve controlled crystal formation, high-pressure homogenization, wet bead milling, and employing supercritical fluids [28]. Other than this the effective approach is encapsulating these compounds within biodegradable nanoparticles. Innovative and biocompatible delivery systems for ellagic acid (EA) currently being developed include pectin-based solid dispersions, cyclodextrin nano sponges, and Supersaturated self-micro emulsifying formulations etc. [26]
2 MATERIALS AND METHOD
ROS Generation in Body;
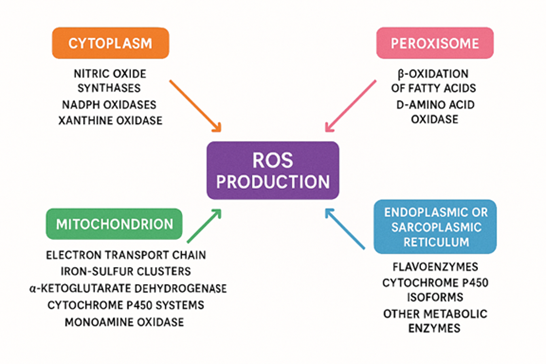
Figure 5. Endogenous formation of unstable oxygen molecular reactive oxygen species (ROS)
The free radicals and Reactive oxygen species (ROS) are generated in different cellular locations, such as the cytoplasm, mitochondria and the endoplasmic or sarcoplasmic reticulum.
Mitochondria serve a crucial role in many aspects such as they not only sustain essential cellular works but also play a crucial function in the onset and progression of many diseases, and the aging process [29]. In mammalian cells, mitochondria are one of the major sources of unstable oxygen molecule or reactive oxygen species, with more than ten distinct sites contributing to ROS formation. This group comprises elements such as the transport electron chain complexes, Iron-sulfur centers, the α-ketoglutarate dehydrogenase complex (α-KGDH), enzymes including mitochondrial cytochrome P450, and monoamine oxidase (MAO).
Apart from mitochondria, reactive oxygen molecules are produced in the cytoplasm, peroxisomes, and the endoplasmic (or sarcoplasmic) reticulum through a variety of enzymatic reactions. Unstable oxygen molecules or ROS are produced in intracellular environment by several key enzyme systems. Among the major contributors are NADPH oxidases, nitric oxide synthases (NOS), xanthine oxidase (XO), and Enzymes interfere in the β-oxidation of fatty acids also play a vital role. In addition, flavoenzymes, cytochrome P450 isoforms, and various metabolic activities affect further to ROS accumulation. Collectively, these sources drive the complex and dynamic production of ROS across different cellular compartments (refer to Figure 3) [30].
ROS Scavenging function of Punicalagin
I. ROS Production:
Punicalagin, at concentration ranging between 1 and 60 µM, significantly reduced reactive oxygen species (ROS) levels in Ellagicacid.hy926 endothelial cells exposed to angiotensin II. This antioxidant effect was validated using flow cytometry and fluorescence-based detection methods [31].
II. Nitric Oxide Production and Punicalagin's Role:
a) Punicalagin Enhances NO2 Production
Punicalagin has been demonstrated to improve nitric oxide production mainly by activating endothelial nitric oxide synthase (eNOS), a mechanism that relies on increased intracellular calcium levels [31,32,33,34].
b) Calcium-Dependent Activation of eNOS
The activation of eNOS is closely controlled by intracellular calcium (Ca²?) levels. An increase in Ca²? allows it to bind with calmodulin, creating a complex that activates eNOS and promotes the production of nitric oxide (NO). This process is one-way, with Ca²? initiating eNOS activation, but not the other way around [35].
c) Mechanisms Regulating eNOS Activation:
After its formation, nitric oxide (NO) enters cells of vascular smooth muscle, where it stimulates miscible guanylate cyclase (sGC), Causing increase in cyclic guanosine monophosphate (cGMP) levels. The increased cGMP activates protein kinase G (PKG), Catalyzing the phosphorylation of eNOS at Ser1177, boosting its activity. This feedback loop helps sustain NO production [32,33,34].
NO → sGC → cGMP → PKG → eNOS phosphorylation → ↑ NO
NO helps maintain eNOS function by reducing oxidative stress. It scavenges oxygen-derived reactive molecules (ROS) particularly superoxide (O?•?), helping preserve tetrahydrobiopterin (BH?), a vital cofactor for eNOS. This prevents eNOS from becoming "uncoupled," a state where it generates superoxide instead of NO [32,33,34]
NO can also influence eNOS expression at the genetic level. It activates transcription factors such as CREB and NF-κB, which may upregulate eNOS gene expression in response to physiological stimuli [32,33,34]. (Figure No. 04)
d) Nitric Oxide and Reactive oxygen molecules (ROS) Reduction
Nitric oxide reacts directly with superoxide radicals to produce peroxynitrite (ONOO?). Although peroxynitrite is itself reactive, this reaction removes harmful superoxide from the system, minimizing oxidative damage [36].
NO negatively regulates NADPH oxidase, a key enzyme in ROS production in both endothelial and immune cells. It inhibits the assembly and activation of NOX isoforms (particularly NOX2 and NOX4), thereby reducing intracellular ROS generation [37].
Through S-nitrosylation, NO can modify cysteine residues on target proteins. This post-translational modification can inhibit enzymes that generate ROS or stabilize antioxidant enzymes, contributing to redox homeostasis and cellular protection [38]. (Figure No. 04)
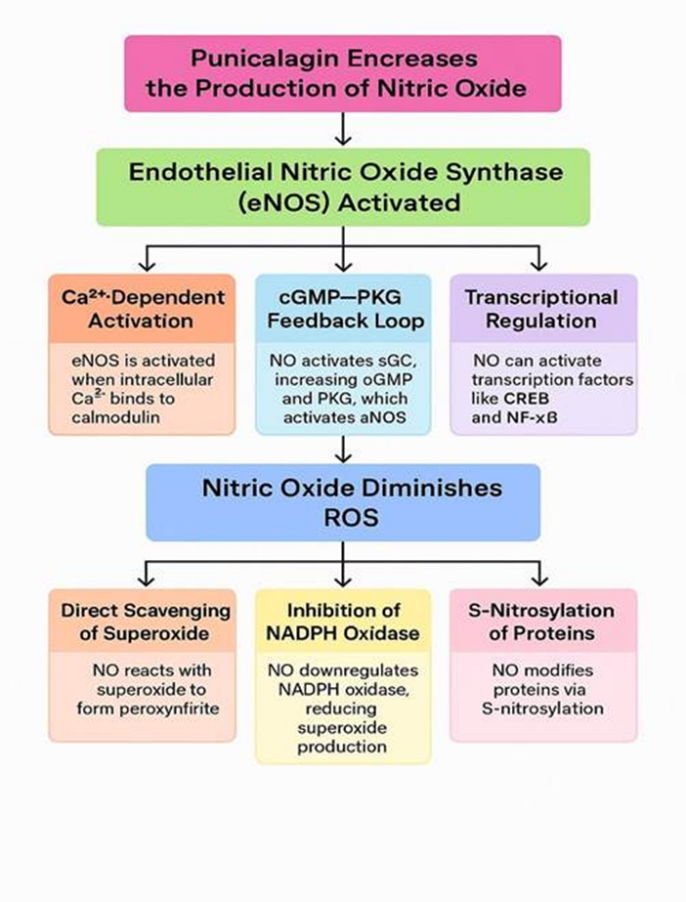
Figure No.5 Punicalagin exerts its scavenging effect by activating endothelial nitric oxide synthase (eNOS), which plays a key role in enhancing nitric oxide production and mitigating oxidative stress.
III. Metal Chelation
Punicalagin can bind transition metal ions like Fe²? and Cu²?, which play crucial roles in Fenton and Haber–Weiss reactions—processes that generate highly reactive hydroxyl radicals (•OH).
By binding and neutralizing these metal ions, punicalagin helps prevent the formation of hydroxyl radicals, thereby reducing metal-induced oxidative damage and protecting cells from oxidative injury.[39]
CONCLUSION
Punicalagin is a bioactive ellagitannin predominantly occur in Punica granatum or pomegranate demonstrates effective antioxidant efficacy through numerous mechanisms. Its potential to directly neutralize unstable oxygen molecules (ROS), suppress NADPH oxidase activity, and bind transition metals like Fe²? and Cu²? makes it an effective regulator of oxidative damage. These actions contribute to decreasing the formation of reactive oxygen species or highly reactive oxygen molecule and lower the overall intracellular ROS levels, thereby maintaining cellular health.
In endothelial cells, punicalagin improve protective effects by enhancing nitric oxide (NO) generation via calcium mediated activation of endothelial nitric oxide synthase (eNOS). Nitric oxide gives antioxidant effect by neutralizing superoxide, preserving eNOS activity via neutralizing the tetrahydrobiopterin (BH?), and influencing eNOS expression Via the activation of signaling pathways like CREB and NF-κB .
Collectively, these actions show that the punicalagin provides therapeutic benefits for diseases related to oxidative stress including the dermal aging, cardiovascular, neurodegenerative, and inflammatory diseases. It has ability to directly diminish ROS and indirectly control redox signaling presents a strong rationale for further drug development research. Future studies focused on its biotransformation of punicalagin, cellular absorption, and potential synergistic effects with other antioxidants will be critical for advancing its medical application
REFERENCES


Azra Banu Gokak, Prajwal Bhosage, A Mechanistic Review on Punicalagin Mediated ROS Scavenging and Cellular Protection, Int. J. of Pharm. Sci., 2025, Vol 3, Issue 9, 2618-2627. https://doi.org/10.5281/zenodo.17182744
 10.5281/zenodo.17182744
10.5281/zenodo.17182744