
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
1Dean Academics, Sai Nath University, Ranchi, Jharkhand 835219, India.
2Associate Professor, Department of Pharmacy, Faculty of Medical Science and Research, Sai Nath University, Ranchi, Jharkhand 835219, India.
3Assistant Professor, Department of Pharmacy, School of Pharmacy, Sai Nath University, Ranchi, Jharkhand 835219, India.
4Student, B. Pharm, Faculty of Medical Science and Research, Sai Nath University, Ranchi, Jharkhand 835219, India.
5Student, B. Pharm, School of Pharmacy, Sai Nath University, Ranchi, Jharkhand 835219, India.
6Assistant Professor of Pharmacology, Department of Pharmacy, Faculty of Medical Science and Research, Sai Nath University, Ranchi, Jharkhand 835219, India
Amphetamine (AMPH) and its derivatives, exhibiting diverse structures and psychoactive effects, share similarities with the botanical diversity of orchids. This review delves into the multifaceted mechanisms underlying their primary biological impact: the elevation of extracellular catecholamines and serotonin levels. We explore these mechanisms independently of classical transmitter release pathways, providing historical context and future research directions. While traditionally referred to in plural form, we cautiously define "the amphetamines" to encompass compounds sharing an ?-methyl-phenethyl-amine motif. Focusing primarily on AMPH and methamphetamine (METH), exemplary catecholamine releasers, we discuss subtle distinctions between these compounds and their differential effects. Furthermore, we examine the historical utilization of natural plant-derived amphetamines, tracing their origins from Ephedra species to contemporary clinical applications. Additionally, we explore the role of trace amines in affective modulation and their potential as endogenous amphetamines. Lastly, we discuss the evolution and impact of synthetic amphetamines, from their inception to contemporary epidemics and clinical uses. Through comprehensive exploration of amphetamine actions on synaptic vesicles and plasma membrane transporters, we unravel their complex pharmacological effects and therapeutic implications
Amphetamine (AMPH) and its various derivatives exhibit a wide range of structural diversity and psychoactive effects, akin to the diversity found in orchids within the plant kingdom [1, 2]. This review explores the multiple mechanisms underlying their primary biological effect: the elevation of extracellular levels of catecholamines and serotonin. This effect occurs independently of the classical mechanisms involved in transmitter release via secretory vesicle fusion [3, 4]. We discuss current concepts within a historical context to provide a framework for future research on this class of drugs, which is both rewarding and addictive, popular yet despised, and capable of both beneficial and destructive effects. Although Alexander Shulgin (1978) argued for the singular use of the term "amphetamine" to denote a specific chemical, linguistic conventions persist in referring to these compounds in the plural form due to the inclusion of the term in the generic names of various compounds sharing the AMPH structure[5, 6]. For the purpose of this review, we define "the amphetamines" cautiously to encompass compounds sharing an ?-methyl-phenethyl-amine motif, as suggested by the generic name of the archetypal compound. Biel and Bopp (1978) outlined the definitive structural features of AMPH, including an unsubstituted phenyl ring, a two-carbon side chain between the phenyl ring and nitrogen, an ?-methyl group, and a primary amino group [7, 8, 9]. The focus of this article is primarily on AMPH and methamphetamine (METH), both exemplary catecholamine releasers with minimal affinity for neuronal receptors, simplifying our review. Although METH deviates from the fourth rule by possessing a secondary amine, it remains one of the most studied compounds in this class [10]. While we do not delve into comparisons of how chemical substitutions alter the efficacy of AMPH-related drugs across various mechanisms, we occasionally reference chain-substituted and ring-substituted amphetamines, as well as non-?-methylated phenethylamines, for comparison. The question often arises regarding the differential effects of AMPH and METH. Studies indicate no significant differences in terms of dopamine release in the striatum, elimination rates, or other pharmacokinetic properties [11, 12]. Human discrimination studies also fail to distinguish between equal doses of the two drugs. However, subtle distinctions exist, such as slightly greater dopamine release by AMPH in the prefrontal cortex, leading to nuanced differences in effects on working memory and behavioral tolerance [13, 14]. AMPH also tends to induce slightly more locomotor activity in rodents than METH, possibly due to indirect effects [15, 16]. Claims suggesting that METH is more addictive, preferred by drug addicts, more potent as a psychostimulant, or exhibits diminished peripheral activity appear unsubstantiated [17, 18]. Traditionally, studies on mechanisms of action focus on AMPH, while studies on neurodegeneration primarily involve METH [19, 20]. This discrepancy may be attributed to the greater availability of METH on the illicit market due to its simpler synthesis methods, which involve either a one-step reduction of ephedrine or pseudoephedrine, or a condensation of phenylacetone and methylamine. The former method yields only the more active S(+)-enantiomer due to the stereochemical purity of ephedrine, a natural product [21, 22, 23].
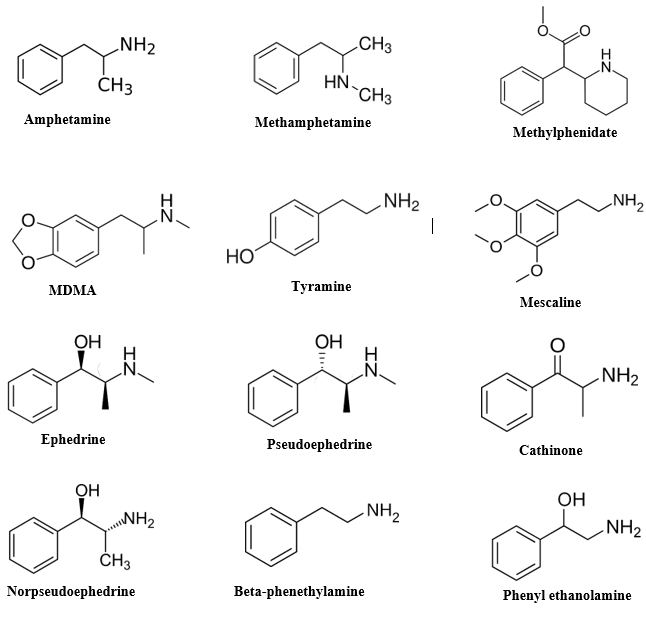
Historical Utilization of Natural Plant-Derived Amphetamines
Similar to nicotine, cocaine, opiates, marijuana, and alcohol, amphetamines have been utilized in their natural plant forms for thousands of years [24, 25, 26]. Ephedra species and the Catha edulis tree, commonly known as khat, have historically been the principal sources of these plant-based stimulants [27, 28]. Ephedra sinica, referred to as ephedra or Ma huang in China, has been unearthed from ancient gravesites in the Middle East and Indian Vedic temples, with controversial associations to soma, a sacred substance in Vedic rituals [29]. Ephedrine, its primary active compound, was identified in the late 19th century. In Western regions, a stimulant tea brewed from a different Ephedra species, Mormon Tea (E. nevadensis), gained popularity, particularly as it was not restricted by the Church of Jesus Christ of Latter-day Saints [30, 31]. However, recent studies suggest that New World Ephedra lacks the alkaloids found in the original ephedrine-containing species [32, 33]. Ephedrine became widely used as an over-the-counter appetite suppressant and performance enhancer, although its dangers were underscored by incidents such as the death of Baltimore Orioles pitcher Steve Bechler in 2003 [34, 35]. Its usage as a dietary supplement was subsequently banned by the FDA in 2004, primarily due to its role as a precursor in the illicit production of methamphetamine [36, 37]. Khat, derived from the leaves of Catha edulis, contains the amphetamines cathinone and cathine [38, 39]. Its use dates back to the 11th century and is prevalent in regions such as Yemen, Kenya, and Somalia [40]. Khat consumption is associated with sociability, euphoria, and appetite suppression [41]. Despite its widespread use, khat is habit-forming and can lead to paranoia and other psychological disturbances [42]. However, withdrawal symptoms are reported to be relatively mild. Khat chewing is deeply embedded in social customs, particularly in Yemen, where dedicated rooms are often set aside for communal khat sessions [43]. While other plant species contain natural amphetamines, including certain Acacia species and Egyptian jasmine, they are not commonly used for administration of these drugs [44, 45]. However, there are plants containing components similar to amphetamines, such as synephrine found in citrus plants and arecoline in the betel nut palm [46, 47]. Mescaline, a phenethylamine analog found in the peyote cactus, has a distinct mechanism of action compared to amphetamines, but has inspired the synthesis of numerous amphetamine derivatives. It has been historically used in Native American rituals and is protected by law for religious purposes [48, 49, 50].
Role of Trace Amines in Affective Modulation and Potential Endogenous Amphetamine Action
It has been long established that decarboxylated metabolites of aromatic amino acids, such as ?-phenethylamine from phenylalanine, phenylethanolamine, tyramine from tyrosine, and tryptamine from tryptophan, are synthesized in both the peripheral nervous system and the brain [51, 52, 53]. These compounds have been implicated in modulating affective behaviors such as excitement and alertness. Interestingly, decreased levels of these "trace amines" have been observed in the urine of depressed patients, while their levels are elevated by marijuana [54].Although trace amines share some mechanisms of action with amphetamines and may potentially act as endogenous amphetamines, they are not stored in substantial quantities in either the central nervous system or the periphery. Exogenous administration of high levels of ?-phenethylamine or its accumulation due to monoamine oxidase (MAO) inhibition can induce psychostimulant responses similar to amphetamines [55, 56]. Like amphetamines, ?-phenethylamine releases dopamine in a manner dependent on the presence of an intact dopamine transporter (DAT) [57]. However, certain behavioral responses to ?-phenethylamine appear to be independent of DAT. The neurotransmitter status of ?-phenethylamine remains uncertain. When present in high concentrations, it is likely to be transported into synaptic vesicles by the vesicular monoamine transporter [58, 59]. However, some studies suggest that ?-phenethylamine may also be released by diffusion across membranes, rather than through reverse transport via catecholamine transporters, and may exhibit insensitivity to reserpine, an inhibitor of vesicular catecholamine uptake [60, 61]. In contrast to amphetamines, trace amines lack the ?-methyl group that inhibits MAO, making them rapidly metabolized in the brain [62]. It is unclear whether local concentrations of ?-phenethylamine in the brain are sufficient to elicit endogenous amphetamine-like effects or activate receptors significantly, although some trace amine compounds show high affinity for recently identified trace amine receptors [63, 64, 65].
The Evolution and Impact of Synthetic Amphetamine: From Invention to Epidemics and Clinical Applications
Synthetic amphetamine (AMPH) was first synthesized in 1887 by Lazar Edeleanu, a Romanian chemist who described its production in his doctoral dissertation under A.W. Hofmann at the University of Berlin [66, 67]. Edeleanu later gained recognition for inventing the method to distill petroleum using sulfur dioxide, which imparted the distinctive odor to gasoline distilleries [68, 69]. AMPH derives its generic name from a contraction of "?-methyl-phenethyl-amine." The rich history of this compound is evident from the 1989 Merck Index, which listed 17 trade names, excluding well-known brands like Adderall, Benzedrine, and Dexedrine, along with numerous nicknames used by drug abusers. Over time, chemists have developed an extensive array of synthetic AMPH derivatives [70, 71]. The concept of sympathomimetic amines, introduced by Barger and Dale, spurred research into various catecholamine-like derivatives for their ability to raise blood pressure and relieve nasal and bronchial congestion [72, 73]. Independently, Gordon Alles resynthesized AMPH in 1927 as part of efforts to develop synthetic sympathomimetics [74]. Walter Hartung and James Munch identified AMPH as a potent sympathomimetic and particularly effective when administered orally. Commercially introduced in 1932 as Benzedrine by the pharmaceutical firm Smith, Kline and French, AMPH gained popularity rapidly. Its first clinical use, for narcolepsy, was reported by Myron Prinzmetal in 1935 [75, 76]. By 1936, Benzedrine tablets became available without prescription, leading to widespread usage, notably among students, artists, musicians, and truck drivers. Initially promoted for over 30 uses, including schizophrenia treatment and seasickness relief, by 1946, AMPH's annual pharmaceutical production reached 10 billion tablets, with a significant portion diverted to the black market [77, 78]. In 1971, legal quotas on AMPH production were imposed by the United States Justice Department. Despite this, AMPH, methamphetamine, and methylphenidate remain widely prescribed for weight control, narcolepsy, and attention deficit disorder [79, 80]. Military use of AMPH for promoting alertness dates back to the Spanish Civil War, with ongoing usage, particularly among fighter pilots [81]. Reports of AMPH abuse and psychosis emerged soon after its introduction, with concerns over addiction becoming more prominent in the mid-1960s. Similar to cocaine, AMPH use tends to occur in epidemic waves, as seen in Japan and Sweden during the mid-20th century [82, 83]. Currently, methamphetamine is the most prevalent illicitly manufactured controlled substance in the United States, with widespread local epidemics reported. MDMA, originally developed for clinical use, remains popular illicitly, despite being assigned Schedule I status in 1985 [84, 85].
Efforts to synthesize AMPH derivatives have led to the exploration of various compounds for clinical applications, including appetite suppression and treatment of Parkinson's Disease [86].
Exploring Multifaceted Sites of Action: A Comprehensive Overview
Early Interplay Between Amphetamine Action and Adrenal Medulla Secretion
Early scientific exploration into the action of amphetamines (AMPH) is intricately tied to the study of the adrenal medulla and its secretion of catecholamines. Catechol, originating from catechu, a plant extract, was traditionally used for dyeing fabric and medicinal purposes [87, 88, 89]. George Oliver, a British physician, observed increased blood pressure upon injecting adrenal gland extract into his son [90, 91]. By 1894, Oliver and Edward Albert Schafer confirmed this effect in dogs [92]. Epinephrine, the active compound, was identified independently by three laboratories in 1897. Its structure was elucidated by Ernst Joseph Friedman in 1906. These discoveries laid the groundwork for the hypothesis of secretory transmission, suggesting that nerves communicate through chemical release [93, 94, 95].
Fig no 2. Mechanism of Action of Amphetamine in Adrenal medulla
Thomas Renton Elliott proposed this idea in 1904 after confirming epinephrine's effects. He also suggested nerve accumulation of epinephrine, though this was demonstrated much later. Pharmacological exploration began with George Barger and Henry H. Dale in 1910, who investigated compounds mimicking epinephrine's effects [96, 97, 98]. They coined the term "sympathomimetic" to describe such compounds. Later research by J.H. Burn and colleagues distinguished between directly and indirectly acting sympathomimetics [99]. Cocaine, derived from Erythoxylon coca, provided insights into physiological mechanisms, particularly as a monoamine uptake transporter inhibitor. Its effects, including its association with Coca-Cola and initial medical enthusiasm, were noted [101, 102]. Alfred Fröhlich and Otto Loewi's work in 1910 on cocaine's interaction with epinephrine added to this understanding [103, 104]. Reserpine, derived from Rauwolfia serpentina, was pivotal in unraveling the actions of sympathomimetics. Its introduction to the West led to significant therapeutic applications, albeit with notable side effects [105, 106]. Arvid Carlsson and others in 1957 showed reserpine's interaction with tyramine, shedding light on sympathomimetic actions [107, 108]. Burn and Rand's 1958 study clarified amphetamines' mechanism by showing their ability to release catecholamines. This explained differences in their actions compared to uptake blockers like cocaine [109, 110]. Later research confirmed amphetamines' role in central dopamine release, emphasizing release over reuptake blockade as the primary mechanism.
Overall, these early investigations laid the foundation for understanding amphetamine action, with subsequent studies refining our knowledge of its physiological effects [111].
Exploring the Role of Plasma Membrane Uptake Transporters and Monoamine Secretory/Synaptic Vesicles in Amphetamine (AMPH) Action: Insights from Reserpine Studies and Genetic Manipulations

Fig no 3 Role of Plasma Membrane Uptake Transporters and Monoamine Secretory/Synaptic Vesicles in Amphetamine (AMPH) Action.
Initial studies using reserpine suggested a role for secretory vesicles in AMPH action. However, subsequent literature presented conflicting findings due to variations in experimental design [112]. Most studies, focusing on dopamine and norepinephrine systems, administered in vivo reserpine injections prior to AMPH administration. These studies generally found that reserpine blocked AMPH-mediated release of norepinephrine, suggesting involvement of vesicular catecholamine [113 ,114, 115]. However, the synthesis of norepinephrine from dopamine within vesicles may affect the interpretation of these results. Combinatorial studies on reserpine and AMPH effects on dopamine release yielded mixed results, with some experiments showing no effect of reserpine on AMPH, while others reported blockade. Attempts to resolve these discrepancies using synaptosomes also produced contradictory results [116, 117]. One explanation for these inconsistencies is that reserpine may upregulate tyrosine hydroxylase activity, leading to increased cytosolic dopamine levels. Shorter exposure to reserpine depleted exocytic dopamine release but did not increase tyrosine hydroxylase activity [118, 119]. This suggests that vesicular catecholamines contribute significantly to AMPH-mediated efflux under typical conditions [120]. Recent studies using genetic manipulations have shed light on the role of synaptic vesicles in AMPH action. Experiments with transfected cell lines and mouse mutants indicate the involvement of both dopamine transporter (DAT) and vesicular monoamine transporter (VMAT) in AMPH-mediated dopamine release [121, 122, 123]. Further investigations using electrochemical detection techniques and mutated transporters support the idea that AMPH acts on both vesicular and plasma membrane transporters, affecting monoamine pools in both synaptic vesicles and the cytosol. Additionally, AMPH-induced de novo dopamine synthesis may contribute to its effects [124, 125, 126]. Furthermore, AMPH affects both vesicular and cytosolic catecholamine pools, with implications for neurotransmitter release and cellular signaling. Its multifaceted actions highlight the complexity of its mechanism of action and its potential therapeutic implications [127, 128]. Amphetamines, including amphetamine and methamphetamine derivatives, exert their pharmacological effects through interactions with monoamine transporters and vesicular monoamine transporters (VMATs) within presynaptic neurons [129, 130]. This review delves into the intricate mechanisms by which amphetamines impact synaptic vesicle dynamics, focusing on neurotransmitter storage, release, and reuptake modulation [131, 132].
The impact on Amphetamines in Synaptic vesicles
Amphetamines are recognized psychostimulants that enhance monoaminergic neurotransmission by modulating monoamine transporters and VMATs. A comprehensive understanding of the interplay between amphetamines and synaptic vesicles is vital for comprehending their pharmacological implications and therapeutic potentiall [133, 134, 135].
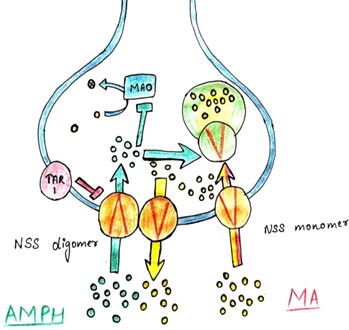
Fig.4. Mechanisms of Amphetamines in Synaptic vesicles
Monoamine Transporter Inhibition
Amphetamines function as substrates for monoamine transporters, such as the dopamine transporter (DAT), norepinephrine transporter (NET), and serotonin transporter (SERT) [136, 137]. By binding to and inhibiting these transporters, amphetamines impede the reuptake of monoamine neurotransmitters from the synaptic cleft into presynaptic neurons, thereby elevating extracellular monoamine levels, notably dopamine and norepinephrine [138, 139, 140].
Vesicular Monoamine Transporter (VMAT) Activation
In addition to monoamine transporter inhibition, amphetamines induce the reverse transport of monoamines by activating VMATs. VMATs play a pivotal role in packaging monoamine neurotransmitters into synaptic vesicles for storage and subsequent release. By facilitating the release of monoamines from synaptic vesicles into the presynaptic neuron's cytoplasm, amphetamines further augment extracellular monoamine concentrations [141, 142 , 143].
Disruption of Vesicular Storage
Extended exposure to high amphetamine concentrations can deplete monoamine stores within synaptic vesicles. This depletion results from VMAT reversal, which prompts the efflux of monoamines from synaptic vesicles into the cytoplasm, leading to a decline in vesicular monoamine content over time. Consequently, neurotransmitter release becomes less reliant on vesicular exocytosis and increasingly dependent on cytoplasmic reserves [144, 145, 146].
Increased Neurotransmitter Release
The cumulative effects of monoamine transporter inhibition and VMAT activation culminate in heightened release of monoamine neurotransmitters, particularly dopamine, into the synaptic cleft. This excessive neurotransmitter release intensifies monoaminergic signaling and contributes to the psychostimulant effects of amphetamines, including heightened arousal, euphoria, and enhanced cognitive function [147, 148, 149]. Lastly, Amphetamines intricately modulate synaptic vesicle function by influencing the dynamics of monoamine neurotransmitter storage, release, and reuptake. A nuanced comprehension of these mechanisms offers valuable insights into the pharmacological effects of amphetamines and holds promise for the development of innovative therapeutic strategies targeting monoaminergic neurotransmission [150, 151].
The impact on Amphetamines in Plasma membrane vesicles
Amphetamines (AMPH) are psychostimulant drugs that exert their pharmacological effects by interacting with plasma membrane transporters involved in the reuptake of monoamine neurotransmitters [152, 153]. This article aims to provide a comprehensive overview of AMPH actions on monoamine transporters, focusing on the dopamine transporter (DAT), norepinephrine transporter (NET), and serotonin transporter (SERT) [154].
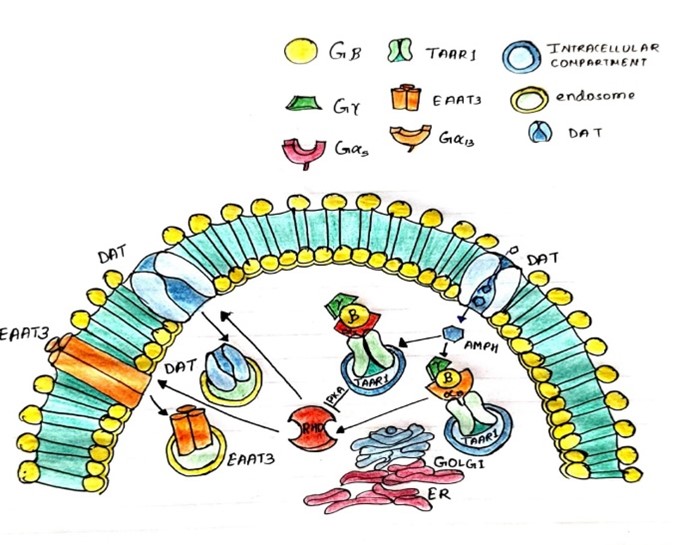
Fig.5. Mechanisms of Amphetamines in Plasma membrane vesicles
Substrate for Monoamine Transporters
AMPH compounds, including amphetamine and methamphetamine, serve as substrates for monoamine transporters. Structurally similar to endogenous neurotransmitters like dopamine, norepinephrine, and serotonin, AMPH molecules are recognized and actively transported by these transporters [155].
Reuptake Inhibition
Upon entering the presynaptic neuron, AMPH competitively inhibits the reuptake of monoamine neurotransmitters from the synaptic cleft into the presynaptic neuron. By binding to and blocking the activity of monoamine transporters, AMPH disrupts the normal reuptake process, leading to an accumulation of neurotransmitters in the synaptic cleft [156].
Enhanced Monoaminergic Signaling
The inhibition of monoamine reuptake by AMPH results in prolonged presence of neurotransmitters in the synaptic cleft, leading to enhanced monoaminergic signaling. This augmentation of neurotransmission contributes to the psychostimulant effects of AMPH, including increased arousal, euphoria, and improved cognitive function [157, 158, 159].
Dopamine Transporter (DAT) Interaction
AMPH exhibits high affinity for DAT, the primary transporter responsible for dopamine reuptake. Binding to DAT, AMPH inhibits dopamine reuptake, leading to elevated extracellular dopamine levels. This increase in dopamine neurotransmission is associated with the rewarding and reinforcing effects of AMPH [160, 161].
Norepinephrine Transporter (NET) Interaction
Similarly, AMPH interacts with NET, the transporter responsible for norepinephrine reuptake. By inhibiting NET activity, AMPH increases extracellular norepinephrine levels, contributing to its stimulant effects on arousal and attention [162].
Serotonin Transporter (SERT) Interaction
While less potent compared to its effects on DAT and NET, AMPH also interacts with SERT, leading to increased extracellular serotonin levels. This modulation of serotonin neurotransmission may contribute to the mood-elevating effects of AMPH [163, 164].
Additional Mechanisms Influencing Extracellular Catecholamine Levels
While amphetamine (AMPH) primarily interacts with plasma membrane transporters, it also influences extracellular catecholamine levels through alternative mechanisms. This section explores these additional pathways, offering insight into the broader regulatory network governing catecholamine neurotransmission [165, 166].
Vesicular Exocytosis and Secretory Vesicles
AMPH's impact extends beyond plasma membrane transporters to include vesicular exocytosis and secretory vesicles. By activating vesicular monoamine transporters (VMATs), AMPH facilitates the release of catecholamines stored within synaptic vesicles into the cytoplasm, a process known as reverse transport, thereby elevating extracellular catecholamine levels [167, 168, 169, 170].
Tyrosine Hydroxylase Activity
AMPH may modulate extracellular catecholamine levels by affecting tyrosine hydroxylase activity, the enzyme critical for catecholamine synthesis. Through enhanced tyrosine hydroxylase activity, AMPH promotes the conversion of tyrosine to L-DOPA, increasing substrate availability for catecholamine synthesis and subsequent release [171].
8Monoamine Oxidase Inhibition
Inhibition of monoamine oxidase (MAO), responsible for catecholamine degradation, represents another potential mechanism. By inhibiting MAO activity, AMPH extends the lifespan of extracellular catecholamines, leading to their accumulation in the synaptic cleft and amplifying neurotransmission [172, 173].
Neuronal Firing and Action Potential Generation
AMPH's stimulatory effects on neuronal firing and action potential generation indirectly influence extracellular catecholamine levels. Increased neuronal excitability promotes neurotransmitter release, including catecholamines, into the synaptic cleft, thereby augmenting extracellular concentrations [174, 175].
Glial Cell Interactions
Evidence suggests that glial cells, particularly astrocytes, play a role in regulating extracellular neurotransmitter levels. AMPH-induced alterations in glial cell function, such as calcium signaling and glutamate release, may indirectly impact catecholamine neurotransmission and contribute to changes in extracellular catecholamine levels [176, 177, 178]. In summary, AMPH's influence on extracellular catecholamine levels encompasses various mechanisms beyond plasma membrane transporters. Through modulation of vesicular exocytosis, tyrosine hydroxylase activity, MAO inhibition, neuronal firing, and glial cell interactions, AMPH exerts complex effects on catecholamine neurotransmission. Understanding these alternative mechanisms provides a comprehensive perspective on AMPH's pharmacological actions and offers potential insights for therapeutic interventions targeting catecholamine dysregulation [179, 180, 181].
CONCLUSION
This article provides a comprehensive exploration of the multifaceted mechanisms underlying the actions of amphetamines, focusing primarily on their impact on neurotransmission. By delving into the historical context, current research, and future directions, it sheds light on the intricate interplay between amphetamines and the central nervous system. The review carefully examines the structural diversity of amphetamines and their psychoactive effects, drawing parallels with the botanical diversity of orchids. It navigates through the elevation of extracellular catecholamines and serotonin levels, elucidating the mechanisms underlying these effects independently of classical transmitter release pathways. The cautious definition of "the amphetamines" encompasses compounds sharing a common structural motif, allowing for a focused discussion primarily on AMPH and methamphetamine. Subtle distinctions between these compounds are explored, alongside their historical utilization from natural plant-derived sources to contemporary clinical applications. Furthermore, the review investigates the role of trace amines in affective modulation and their potential as endogenous amphetamines, providing insights into the broader neurobiological landscape. It also traces the evolution and impact of synthetic amphetamines, from their inception to contemporary epidemics and clinical uses. Through a comprehensive exploration of amphetamine actions on synaptic vesicles and plasma membrane transporters, the article unravels their complex pharmacological effects and therapeutic implications. By dissecting the mechanisms underlying amphetamine-mediated neurotransmission, it offers valuable insights into both the rewarding and addictive nature of these compounds, as well as their potential therapeutic applications. Overall, this review contributes to our understanding of the multifaceted mechanisms of amphetamines and their impact on neurotransmission, paving the way for future research and clinical interventions in this field.
REFERENCES:













K. Rajeswar Dutt, Mahesh Kumar Yadav, Abrarul Haque, Maheshwar Kumar, Abhishek Kumar, Sunny Kumar, Deepak Kumar, Pratik Mondal, Nisha Kumari, Balraj Kumar, Gautam Mahto, Arnab Roy, Exploring The Multifaceted Mechanisms Of Amphetamines And Their Impact On Neurotransmission, Int. J. of Pharm. Sci., 2024, Vol 2, Issue 5, 307-328. https://doi.org/10.5281/zenodo.11142830
 10.5281/zenodo.11142830
10.5281/zenodo.11142830