
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
1Assistant Professor, Gawande College of Pharmacy, Sakharkherda, Buldhana
2Department of Pharmacology, Institute of Pharmaceutical Education and Research, Borgaon meghe, Wardha
Neurological disorders, including Alzheimer’s disease, Parkinson’s disease, glioblastoma, ischemic stroke, multiple sclerosis, amyotrophic lateral sclerosis, Huntington’s disease, traumatic brain injury, and spinal cord injury, remain major causes of mortality and long-term disability worldwide. Despite significant advances in neuroscience and pharmacotherapy, the clinical management of these disorders is severely limited by the complex pathophysiology of the central nervous system (CNS), inadequate regeneration of neural tissue, poor drug stability, systemic toxicity, and the restrictive nature of the blood–brain barrier (BBB), which substantially limits the delivery of therapeutic agents to diseased brain regions. In recent years, engineered exosomes have emerged as highly promising biological nanocarriers owing to their intrinsic biocompatibility, nanoscale dimensions, low immunogenicity, prolonged circulation, and natural ability to cross the BBB. In addition to serving as efficient drug delivery systems, exosomes have gained considerable attention as minimally invasive biomarkers because their molecular cargo reflects the physiological and pathological status of their parent cells. Advances in genetic engineering, surface functionalization, cargo-loading technologies, and synthetic biology have further enhanced the therapeutic potential of engineered exosomes by improving targeting specificity, drug-loading efficiency, and controlled intracellular release. Preclinical investigations have demonstrated encouraging outcomes in multiple neurological disorders through the delivery of small-molecule drugs, proteins, peptides, messenger RNA, microRNA, small interfering RNA, CRISPR/Cas gene-editing components, and neuroprotective biomolecules. Nevertheless, important challenges, including manufacturing standardization, large-scale production, product heterogeneity, quality control long-term safety, pharmacokinetics, regulatory approval, and clinical translation, continue to impede widespread clinical application. This review critically examines recent advances in engineered exosomes for neurological disorders, emphasizing their roles as diagnostic biomarkers, brain-targeted drug delivery systems, and emerging therapeutic platforms while discussing current translational challenges, future technological innovations, and opportunities for precision neurology
1.1 Global Burden of Neurological Disorders
Neurological disorders are a major global health challenge, contributing significantly to mortality, disability, and healthcare costs. The growing burden is driven by population ageing, longer life expectancy, and improved diagnosis. Disorders such as Alzheimer’s disease (AD), Parkinson’s disease (PD), ischemic stroke, glioblastoma, multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), Huntington’s disease (HD), traumatic brain injury (TBI), and spinal cord injury (SCI) have limited disease-modifying treatments despite advances in neuroscience. Their progression is associated with interconnected pathological mechanisms, including oxidative stress, mitochondrial dysfunction, chronic neuroinflammation, excitotoxicity, impaired autophagy, abnormal protein aggregation, synaptic dysfunction, apoptosis, and neuronal loss, highlighting the need for multifunctional therapeutic strategies capable of targeting multiple pathways within the central nervous system (CNS)1.
1.2 Limitations of Conventional Therapies
Conventional therapies for neurological disorders face major challenges, primarily due to the blood–brain barrier (BBB), which prevents most drugs from reaching the central nervous system (CNS). Nearly 98% of small-molecule drugs and almost all biological therapeutics cannot effectively cross the BBB. In addition, conventional drug delivery is often limited by rapid clearance, poor bioavailability, low target specificity, enzymatic degradation, and systemic toxicity. Although nanoparticle-based systems have improved drug delivery, they still face issues such as immunogenicity, poor biodegradability, and limited targeting efficiency, creating a need for safer and more effective delivery platforms
1.3 Biology and Characteristics of Exosomes
Exosomes are naturally occurring extracellular vesicles (30–150 nm) released by almost all mammalian cells through the endosomal pathway. They carry diverse bioactive molecules, including proteins, lipids, mRNAs, microRNAs, long non-coding RNAs, and other signaling molecules that reflect the condition of their parent cells. Unlike synthetic nanocarriers, exosomes have a natural lipid bilayer that protects their cargo, low immunogenicity, excellent biocompatibility, prolonged circulation, and the ability to cross the blood–brain barrier (BBB). These unique properties make exosomes promising candidates for targeted drug delivery and intercellular communication in neurological disorders
1.4 Evolution of Engineered Exosomes
The rapid expansion of extracellular vesicle research has transformed exosomes from passive mediators of intercellular communication into highly sophisticated therapeutic nanoplatforms. Early investigations primarily focused on understanding exosome biogenesis and physiological functions; however, recent advances in molecular engineering have enabled extensive modification of exosomes to improve their therapeutic performance. Engineered exosomes can now be generated through genetic modification of donor cells, chemical conjugation of membrane ligands, membrane hybridization, biomimetic engineering, and advanced cargo-loading technologies. These approaches have significantly enhanced targeting specificity, drug-loading efficiency, intracellular delivery, and controlled release while preserving the favorable biological characteristics of native exosomes.
The ability to encapsulate diverse therapeutic molecules—including small-molecule drugs, proteins, peptides, messenger RNAs, microRNAs, small interfering RNAs, and genome-editing systems—has expanded the potential applications of engineered exosomes across numerous neurological diseases. In parallel, improvements in manufacturing technologies, purification methods, and characterization techniques have accelerated progress toward clinical translation2.
1.5 Rationale for Brain-Targeted Drug Delivery
Brain-targeted drug delivery remains a major challenge because the blood–brain barrier (BBB) restricts the entry of most therapeutic agents into the central nervous system (CNS). Engineered exosomes offer a promising solution due to their natural ability to cross the BBB, excellent biocompatibility, and low immunogenicity. Surface modification with targeting ligands such as rabies virus glycoprotein (RVG) peptide, Angiopep-2, transferrin, apolipoprotein E (ApoE), and cyclic arginine–glycine–aspartate (cRGD) peptides further enhances receptor-mediated transport and selective delivery to neuronal and glial cells, making engineered exosomes attractive carriers for precision therapy in neurological disorders3
Table No. 1 Comparison Between Native and Engineered Exosomes
|
Feature |
Native Exosomes |
Engineered Exosomes |
|
Source |
Naturally secreted by cells |
Naturally secreted but deliberately modified |
|
Cargo |
Endogenous proteins, lipids, and nucleic acids |
Therapeutic drugs, miRNA, siRNA, mRNA, proteins, CRISPR components |
|
Targeting Ability |
Limited, non-specific |
Enhanced through surface ligands or genetic engineering |
|
BBB Penetration |
Moderate |
Improved with targeting modifications |
|
Therapeutic Efficacy |
Natural biological activity |
Higher efficacy and precision delivery |
|
Drug Loading |
Limited natural loading |
Active loading by electroporation, incubation, sonication, etc. |
|
Clinical Application |
Biomarkers and natural therapeutics |
Targeted drug and gene delivery for neurological diseases |
|
Challenges |
Low targeting specificity |
Manufacturing complexity, scalability, and regulatory approval |
1.6 Scope and Objectives of the Review
This review critically summarizes recent advances in the use of engineered exosomes for neurological disorders. It focuses on exosome biology, engineering strategies, biomarker discovery, blood–brain barrier (BBB) transport, targeted drug delivery, therapeutic applications, clinical translation, manufacturing challenges, regulatory aspects, and future perspectives. By integrating findings from recent preclinical and clinical studies, this review provides an overview of the potential of engineered exosomes to improve the diagnosis and treatment of neurological disorders through precision nanomedicine.
2. BIOLOGY AND BIOGENESIS OF EXOSOMES
2.1 Biology and Biogenesis of Exosomes
Exosomes are 30–150 nm extracellular vesicles (EVs) secreted by almost all eukaryotic cells and play a vital role in intercellular communication by transporting proteins, lipids, RNAs, and other bioactive molecules. In the central nervous system (CNS), neurons, astrocytes, oligodendrocytes, microglia, endothelial cells, and neural stem cells release exosomes that regulate neuronal communication, synaptic plasticity, immune responses, and tissue repair. Their molecular cargo reflects the physiological or pathological state of the parent cell, making them valuable as both biomarkers and therapeutic carriers2.
Exosome biogenesis begins with the inward budding of the plasma membrane to form early endosomes, which mature into late endosomes containing multivesicular bodies (MVBs). During this process, intraluminal vesicles (ILVs) are generated through inward budding of the endosomal membrane. MVBs either fuse with lysosomes for degradation or with the plasma membrane to release ILVs as mature exosomes. This process is regulated by the endosomal sorting complex required for transport (ESCRT) machinery, Rab GTPases, and SNARE proteins, which control cargo sorting, vesicle trafficking, and exosome secretion4.
Within the CNS, exosomes maintain neuronal homeostasis by mediating communication between neurons and glial cells. Neuronal exosomes support synaptic function and neuronal survival, astrocyte-derived exosomes regulate metabolism and antioxidant defense, oligodendrocyte-derived exosomes maintain myelin integrity, and microglial exosomes modulate neuroinflammatory responses. However, under pathological conditions, exosomes can also facilitate the spread of disease-associated proteins such as amyloid-β, tau, and α-synuclein, contributing to the progression of neurodegenerative diseases. Conversely, stem cell-derived exosomes enriched with neuroprotective proteins and microRNAs exhibit anti-inflammatory and regenerative effects, highlighting their therapeutic potential2.
The natural origin of exosomes provides several advantages over synthetic nanocarriers, including low immunogenicity, excellent biocompatibility, protection of therapeutic cargo, prolonged circulation, and intrinsic targeting ability. Their capacity to cross the blood–brain barrier (BBB) and deliver bioactive molecules to specific CNS cells has made engineered exosomes promising platforms for brain-targeted drug delivery and regenerative medicine4.
2.2 ESCRT-Dependent Pathway
The endosomal sorting complex required for transport (ESCRT) is the best-characterized mechanism regulating exosome biogenesis and cargo sorting. It consists of four protein complexes (ESCRT-0, ESCRT-I, ESCRT-II, and ESCRT-III) together with accessory proteins such as apoptosis-linked gene 2-interacting protein X (ALIX), tumor susceptibility gene 101 (TSG101), and vacuolar protein sorting-associated protein 4 (VPS4). These components sequentially mediate cargo recognition, membrane budding, intraluminal vesicle (ILV) formation, membrane scission, and exosome release.
Exosome biogenesis begins when ESCRT-0, mainly through HRS and STAM, recognizes ubiquitinated proteins and recruits them to endosomal membranes. ESCRT-I and ESCRT-II then induce inward membrane budding and selectively package proteins, lipids, messenger RNAs (mRNAs), microRNAs (miRNAs), and other biomolecules into developing ILVs. Finally, ESCRT-III drives membrane constriction and scission to generate mature ILVs, while VPS4 disassembles and recycles ESCRT components for subsequent rounds of vesicle formation.
In the central nervous system (CNS), the ESCRT pathway is essential for neuronal communication, synaptic plasticity, protein homeostasis, and intracellular trafficking. Dysregulation of ESCRT proteins impairs protein degradation and lysosomal function, promoting the accumulation of pathogenic proteins associated with Alzheimer’s disease, Parkinson’s disease, amyotrophic lateral sclerosis, and other neurodegenerative disorders. Because ESCRT proteins regulate both cargo selection and exosome secretion, they have become important targets for engineering exosomes with improved therapeutic cargo loading and brain-targeting efficiency. Although ESCRT-dependent biogenesis is the predominant mechanism, exosomes can also be generated through ESCRT-independent pathways, highlighting the complexity of exosome formation5.
2.3 ESCRT-Independent Pathway
In addition to the endosomal sorting complex required for transport (ESCRT) machinery, exosomes can also be generated through ESCRT-independent pathways. These mechanisms rely primarily on ceramide-mediated membrane remodeling, tetraspanin-enriched microdomains, and lipid–protein interactions to promote intraluminal vesicle (ILV) formation and cargo sorting. The persistence of exosome secretion after ESCRT inhibition demonstrates that these alternative pathways play an important role in exosome biogenesis.
The ceramide-dependent pathway is the best-characterized ESCRT-independent mechanism. Ceramide, produced by neutral sphingomyelinase (nSMase) through sphingomyelin hydrolysis, induces negative membrane curvature that promotes inward budding of the endosomal membrane and ILV formation. Inhibition of nSMase significantly reduces exosome secretion, confirming the critical role of ceramide in this process. In addition, sphingolipid metabolites such as sphingosine-1-phosphate (S1P) regulate multivesicular body maturation, intracellular trafficking, and vesicle release through multiple signaling pathways.
Another important mechanism involves tetraspanins, particularly CD9, CD63, CD81, and CD82, which organize specialized membrane microdomains that facilitate membrane budding, cargo clustering, and exosome formation. These proteins also regulate interactions with integrins, adhesion molecules, and signaling receptors, contributing to the selective incorporation of proteins and nucleic acids into exosomes. Furthermore, exosomal membranes are enriched in cholesterol, sphingomyelin, phosphatidylserine, and glycosphingolipids, forming lipid raft domains that enhance membrane stability and support efficient vesicle formation6.
Within the central nervous system (CNS), ESCRT-independent pathways contribute to neuronal communication, synaptic plasticity, myelin maintenance, and neuroimmune regulation. However, altered lipid metabolism and tetraspanin expression can promote the spread of pathogenic proteins, including amyloid-β, tau, and α-synuclein, during neurodegenerative diseases. Conversely, stem cell-derived exosomes generated through these pathways often carry neuroprotective proteins and regulatory microRNAs that reduce neuroinflammation and promote tissue repair. Understanding ESCRT-independent mechanisms therefore provides valuable opportunities to optimize exosome engineering, improve cargo loading, and enhance brain-targeted drug delivery6.
2.4 Cargo Sorting Mechanisms in Exosomes
The biological function of exosomes is determined by their selectively packaged cargo, including proteins, lipids, messenger RNAs (mRNAs), microRNAs (miRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), DNA fragments, metabolites, and signaling molecules. Cargo loading is a highly regulated process rather than random encapsulation, allowing donor cells to transfer specific biological signals to recipient cells. This selective packaging is essential for intercellular communication, immune regulation, neuronal homeostasis, and disease progression.
Cargo sorting occurs during the maturation of multivesicular bodies (MVBs) through both ESCRT-dependent and ESCRT-independent pathways. In the ESCRT-dependent mechanism, proteins such as apoptosis-linked gene 2-interacting protein X (ALIX), tumor susceptibility gene 101 (TSG101), syntenin, and syndecans regulate the selective incorporation of membrane proteins into intraluminal vesicles (ILVs). In contrast, ESCRT-independent sorting involves tetraspanins (CD9, CD63, CD81, and CD82) and lipid raft microdomains, which facilitate membrane organization and cargo enrichment.
Selective RNA loading is mediated by RNA-binding proteins, including heterogeneous nuclear ribonucleoproteins (hnRNPs), Argonaute 2 (AGO2), Y-box-binding protein 1 (YBX1), and SYNCRIP, which recognize specific RNA motifs and direct mRNAs, miRNAs, lncRNAs, and circRNAs into exosomes. Among these, exosomal miRNAs are particularly important because they regulate neuronal survival, synaptic plasticity, neuroinflammation, apoptosis, and angiogenesis. Disease-specific changes in exosomal miRNA profiles have been reported in Alzheimer’s disease, Parkinson’s disease, ischemic stroke, glioblastoma, multiple sclerosis, amyotrophic lateral sclerosis, and Huntington’s disease, highlighting their value as diagnostic biomarkers and therapeutic targets7.
Exosomal membranes are enriched with cholesterol, ceramide, sphingomyelin, phosphatidylserine, and glycosphingolipids, which maintain membrane stability, promote vesicle fusion, and influence biodistribution and cellular uptake. Environmental factors such as hypoxia, oxidative stress, inflammation, and metabolic alterations further modify cargo composition by regulating intracellular signaling pathways. These dynamic changes generate disease-specific molecular signatures that reflect the physiological status of donor cells.
A detailed understanding of cargo sorting mechanisms has enabled the development of engineered exosomes for targeted drug delivery. Endogenous approaches genetically modify donor cells to enrich therapeutic proteins or nucleic acids within secreted exosomes, whereas exogenous methods—including electroporation, sonication, extrusion, passive incubation, freeze–thaw cycling, and microfluidic loading—allow incorporation of therapeutic molecules after exosome isolation. These strategies have significantly improved the delivery of small-molecule drugs, proteins, peptides, mRNAs, miRNAs, siRNAs, antisense oligonucleotides, and CRISPR/Cas gene-editing systems, supporting the development of precision nanomedicine for neurological disorders8.
2.5 Molecular Composition of Exosomes
Exosomes possess a highly organized molecular composition that reflects the physiological state and cellular origin of their parent cells. Their lipid bilayer encloses a diverse cargo of proteins, lipids, messenger RNAs (mRNAs), microRNAs (miRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), DNA fragments, metabolites, and signaling molecules, which collectively regulate intercellular communication and determine their therapeutic potential.
The exosomal membrane is enriched with cholesterol, sphingomyelin, ceramide, phosphatidylserine, phosphatidylcholine, phosphatidylethanolamine, and glycosphingolipids. These lipids maintain membrane stability, protect encapsulated cargo from degradation, and facilitate membrane fusion and cellular uptake. Lipid raft microdomains also contribute to exosome biodistribution, signaling, and immune regulation.
Proteins are major functional components of exosomes. Common exosomal markers include CD9, CD63, CD81, CD82, apoptosis-linked gene 2-interacting protein X (ALIX), tumor susceptibility gene 101 (TSG101), heat shock proteins (HSP70 and HSP90), annexins, flotillins, and Rab GTPases, which regulate cargo sorting, vesicle trafficking, membrane organization, and cell recognition. Integrins, adhesion molecules, and major histocompatibility complex (MHC) proteins further influence tissue-specific targeting and immune responses.
Exosomes also transport functional nucleic acids, including mRNAs, miRNAs, lncRNAs, circRNAs, transfer RNAs (tRNAs), ribosomal RNAs (rRNAs), PIWI-interacting RNAs (piRNAs), and DNA fragments. Among these, miRNAs are particularly important because they regulate neuronal differentiation, synaptic plasticity, angiogenesis, neuroinflammation, apoptosis, and oxidative stress. Disease-specific alterations in exosomal RNA profiles have been reported in Alzheimer’s disease, Parkinson’s disease, glioblastoma, ischemic stroke, multiple sclerosis, amyotrophic lateral sclerosis, and Huntington’s disease, supporting their use as diagnostic biomarkers and therapeutic targets9.
The molecular composition of exosomes varies according to cell type and pathological conditions. Neuron-derived exosomes are enriched in synaptic proteins, astrocyte-derived exosomes contain neurotrophic and antioxidant molecules, oligodendrocyte-derived exosomes transport myelin-associated proteins, whereas mesenchymal stem cell-derived exosomes are enriched with regenerative proteins, growth factors, and anti-inflammatory miRNAs. These unique molecular signatures determine the biological function and therapeutic potential of exosomes.
Comprehensive characterization of exosomal composition is essential for both research and clinical applications. Techniques such as transmission electron microscopy (TEM), cryogenic electron microscopy (cryo-EM), nanoparticle tracking analysis (NTA), flow cytometry, western blotting, mass spectrometry-based proteomics, lipidomics, and next-generation sequencing (NGS) are widely used to evaluate exosome morphology, size, concentration, and molecular cargo. Detailed molecular profiling has provided the foundation for engineering exosomes with improved targeting specificity, enhanced drug-loading capacity, and greater therapeutic efficacy for neurological disorders9.
2.6 Cellular Uptake and Intercellular Communication
The biological activity of exosomes depends on their ability to recognize, bind, and deliver functional cargo to recipient cells. Through the transfer of proteins, lipids, messenger RNAs (mRNAs), microRNAs (miRNAs), and other bioactive molecules, exosomes regulate intercellular communication, gene expression, immune responses, and tissue homeostasis. In the central nervous system (CNS), they mediate communication among neurons, astrocytes, oligodendrocytes, microglia, endothelial cells, and neural stem cells, thereby maintaining neuronal function and supporting tissue repair.
Cellular uptake begins with the interaction of exosomal surface molecules, including tetraspanins, integrins, immunoglobulin superfamily proteins, proteoglycans, and glycoproteins, with complementary receptors on recipient cells. These ligand–receptor interactions determine tissue specificity and promote selective uptake. Exosomes are internalized through multiple pathways, including clathrin-mediated endocytosis, caveolin-mediated endocytosis, macropinocytosis, phagocytosis, and direct membrane fusion. The predominant mechanism depends on the cell type, receptor expression, and exosomal membrane composition.
Following internalization, exosomes either fuse with endosomal membranes to release their cargo into the cytoplasm or undergo lysosomal degradation. Direct membrane fusion enables rapid cytoplasmic delivery of therapeutic molecules, including small interfering RNAs (siRNAs), messenger RNAs (mRNAs), proteins, and CRISPR/Cas gene-editing systems, thereby avoiding lysosomal degradation. In some cases, exosomes also activate intracellular signaling through surface receptor interactions without complete internalization.
Within the CNS, exosome-mediated communication regulates synaptic plasticity, neuronal survival, myelin maintenance, angiogenesis, neuroinflammation, and immune homeostasis. Neuron-derived exosomes transport synaptic proteins and regulatory miRNAs, astrocyte-derived exosomes provide neurotrophic and antioxidant support, oligodendrocyte-derived exosomes maintain axonal integrity, and microglial exosomes modulate inflammatory responses. Under pathological conditions, exosomes can facilitate the spread of amyloid-β, hyperphosphorylated tau, and α-synuclein, contributing to the progression of Alzheimer’s disease and Parkinson’s disease. Conversely, mesenchymal stem cell-derived exosomes enriched with neuroprotective proteins and miRNAs reduce neuroinflammation, promote angiogenesis, and enhance functional recovery after neurological injury.
Understanding cellular uptake mechanisms has enabled the development of engineered exosomes with improved targeting efficiency. Surface modification using peptides, antibodies, aptamers, and receptor-specific ligands enhances selective uptake by diseased tissues while minimizing off-target effects. These strategies improve intracellular cargo delivery and support the use of engineered exosomes as effective brain-targeted nanocarriers for neurological disorders10.
2.7 Biological Functions of Exosomes in the Central Nervous System
Exosomes are key mediators of intercellular communication in the central nervous system (CNS), facilitating the exchange of proteins, lipids, messenger RNAs (mRNAs), microRNAs (miRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), and signaling molecules among neurons, astrocytes, oligodendrocytes, microglia, endothelial cells, and neural stem cells. By protecting and transporting these bioactive molecules, exosomes regulate neuronal homeostasis, synaptic function, immune responses, and tissue repair.
During brain development, exosomes promote neurogenesis, neuronal differentiation, axonal growth, and synapse formation by modulating signaling pathways such as Wnt/β-catenin, Notch, phosphoinositide 3-kinase/protein kinase B (PI3K/Akt), mitogen-activated protein kinase (MAPK), and transforming growth factor-β (TGF-β). Neuron-derived exosomes also regulate synaptic plasticity, neurotransmitter release, dendritic spine remodeling, learning, and memory through the transfer of synaptic proteins and regulatory miRNAs.
Glial cell-derived exosomes play equally important roles in maintaining CNS homeostasis. Astrocyte-derived exosomes deliver neurotrophic factors, antioxidant enzymes, and metabolic regulators that support neuronal survival and protect against oxidative stress. Oligodendrocyte-derived exosomes transport myelin-associated proteins and metabolic enzymes required for axonal maintenance and myelin integrity, whereas microglial exosomes regulate immune surveillance, synaptic pruning, and neuroinflammatory responses by transferring cytokines, chemokines, and immune-regulatory miRNAs.
Exosomes also contribute to blood–brain barrier (BBB) integrity and neurovascular communication. Endothelial cell-derived exosomes regulate angiogenesis, vascular remodeling, and endothelial function, while astrocyte-derived exosomes enhance tight junction integrity and maintain BBB homeostasis. These interactions are essential for normal brain function and become increasingly important during ischemic injury and neuroinflammation.
In neurological disorders, exosome-mediated communication can either promote disease progression or support tissue repair. Exosomes carrying amyloid-β, hyperphosphorylated tau, and α-synuclein contribute to the spread of pathological proteins in Alzheimer’s disease and Parkinson’s disease, whereas glioblastoma-derived exosomes facilitate tumor growth, angiogenesis, immune evasion, and therapeutic resistance. In contrast, mesenchymal stem cell-derived exosomes exhibit potent neuroprotective effects by reducing neuroinflammation, inhibiting apoptosis, promoting angiogenesis, enhancing neurogenesis, and stimulating functional recovery after ischemic stroke, traumatic brain injury, spinal cord injury, and other neurodegenerative disorders.
The natural ability of exosomes to cross the BBB, combined with their low immunogenicity and high biocompatibility, has made them attractive therapeutic platforms. Engineering approaches that modify exosomal cargo or decorate the membrane with targeting ligands further enhance their capacity for precise brain-targeted drug delivery, highlighting their growing importance in regenerative neurology and precision nanomedicine11.
3. EXOSOMES AS BIOMARKERS IN NEUROLOGICAL DISORDERS
The identification of reliable biomarkers for neurological disorders remains a major challenge because many central nervous system (CNS) diseases progress silently before clinical symptoms become apparent. Conventional diagnostic methods, including neuroimaging, cerebrospinal fluid (CSF) analysis, electrophysiological testing, and neurological examinations, often have limited sensitivity for early diagnosis, while repeated CSF sampling is invasive. Exosomes have emerged as promising liquid biopsy biomarkers because they are secreted by most CNS cells, can cross the blood–brain barrier (BBB), and are detectable in accessible body fluids such as plasma, serum, cerebrospinal fluid, saliva, and urine. Their molecular cargo closely reflects the physiological and pathological state of their parent cells, making them valuable for early diagnosis, disease monitoring, and prognosis.
The phospholipid bilayer of exosomes protects proteins, messenger RNAs (mRNAs), microRNAs (miRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), lipids, and metabolites from degradation, ensuring high molecular stability during circulation. Advances in immunoaffinity isolation techniques also enable selective enrichment of neuron-, astrocyte-, oligodendrocyte-, endothelial-, and microglia-derived exosomes, improving the specificity of brain-derived biomarkers from peripheral blood. Consequently, exosome-based liquid biopsy offers a minimally invasive approach for disease stratification, therapeutic monitoring, and personalized management of neurological disorders.
Exosomal cargo changes dynamically during disease progression, providing important insights into disease mechanisms. Distinct alterations in exosomal proteins, lipids, cytokines, and regulatory RNAs have been reported in neurodegenerative, cerebrovascular, neuroinflammatory, and brain tumor disorders. Furthermore, the integration of proteomics, transcriptomics, lipidomics, metabolomics, next-generation sequencing, and machine learning is accelerating the discovery of highly sensitive and disease-specific exosomal biomarkers, supporting the development of precision neurology and personalized medicine12.
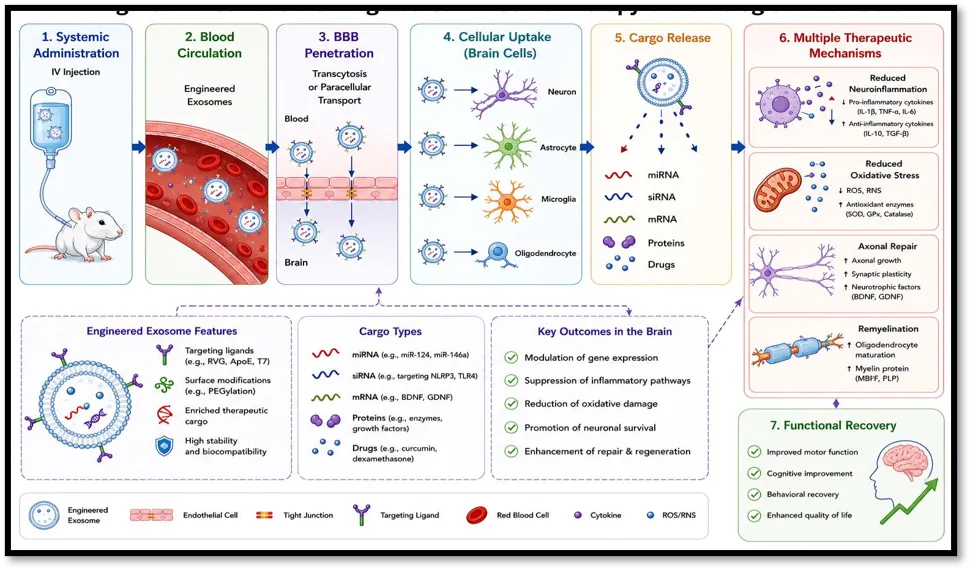
Fig No. 1 Mechanism of Engineered Exosome Therapy in Neurological Disorders
3.1 Exosomal Biomarkers in Alzheimer's Disease
Alzheimer's disease (AD) is the leading cause of dementia and is characterized by amyloid-β (Aβ) plaque deposition, hyperphosphorylated tau accumulation, synaptic dysfunction, neuroinflammation, mitochondrial dysfunction, and progressive neuronal loss. Although neuroimaging and cerebrospinal fluid (CSF) biomarkers have improved diagnosis, their high cost and invasive nature limit routine monitoring. Exosomes have emerged as promising liquid biopsy biomarkers because neuron-derived exosomes cross the blood–brain barrier (BBB) and carry disease-specific molecular cargo that reflects early pathological changes.
Among exosomal protein biomarkers, Aβ42, total tau, phosphorylated tau, neurofilament light chain (NfL), synaptophysin, neurogranin, and growth-associated protein 43 (GAP-43) have shown strong diagnostic and prognostic value. Elevated levels of Aβ42 and phosphorylated tau in neuron-derived exosomes are associated with cognitive decline and progression from mild cognitive impairment (MCI) to AD. In addition, dysregulated exosomal microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs) regulate amyloid processing, tau phosphorylation, oxidative stress, and neuroinflammation, making them promising biomarkers for early diagnosis and disease monitoring. Integration of multi-omics approaches is expected to further improve diagnostic accuracy. However, standardization of exosome isolation, characterization, and analytical methods remains essential for successful clinical translation13.
3.2 Exosomal Biomarkers in Parkinson's Disease
Parkinson's disease (PD) is the second most common neurodegenerative disorder and is characterized by the progressive loss of dopaminergic neurons and intracellular aggregation of α-synuclein into Lewy bodies. Because neurodegeneration begins years before motor symptoms appear, reliable early biomarkers are urgently needed. Exosomes are attractive candidates because neuron-derived vesicles cross the blood–brain barrier (BBB) and carry disease-specific molecular signatures.
Among exosomal proteins, total α-synuclein, oligomeric α-synuclein, phosphorylated α-synuclein, tau protein, amyloid-β1–42, DJ-1, leucine-rich repeat kinase 2 (LRRK2), and inflammatory proteins have demonstrated diagnostic value. Exosomal α-synuclein is the most extensively studied biomarker and is closely associated with disease severity and progression. In addition, dysregulated exosomal miRNAs, including miR-24, miR-23b-3p, miR-195-3p, miR-29c, miR-331-5p, miR-21, miR-1, and miR-22, regulate pathways related to autophagy, mitochondrial dysfunction, oxidative stress, neuroinflammation, and neuronal survival. lncRNAs and circRNAs further complement these biomarkers by providing additional information on gene regulation and disease mechanisms.
Recent advances in proteomics, transcriptomics, lipidomics, metabolomics, and next-generation sequencing have enabled comprehensive profiling of exosomal cargo, improving biomarker sensitivity and specificity. Blood-derived neuron-specific exosomes also support repeated, minimally invasive monitoring of disease progression and therapeutic response. Nevertheless, variability in isolation techniques, analytical platforms, and patient heterogeneity remains a major challenge. Future clinical application will require standardized protocols and validation in large multicenter studies14.
3.3 Exosomal Biomarkers in Glioblastoma
Glioblastoma (GBM) is the most aggressive primary malignant tumor of the central nervous system and remains associated with poor survival despite advances in surgery, radiotherapy, and temozolomide chemotherapy. Early diagnosis and continuous disease monitoring are challenging because conventional tissue biopsy is invasive and cannot adequately capture tumor heterogeneity. Exosomes have emerged as promising liquid biopsy biomarkers because they are released by glioblastoma cells, cross the blood–brain barrier, and preserve the molecular characteristics of their parent tumor cells15.
Glioblastoma-derived exosomes carry proteins, messenger RNAs, microRNAs (miRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), DNA fragments, lipids, and metabolites that regulate tumor proliferation, angiogenesis, invasion, immune evasion, and therapeutic resistance. Protein biomarkers such as EGFR/EGFRvIII, PD-L1, VEGF, MMP-2, MMP-9, TGF-β, integrins, and CD44, together with dysregulated miRNAs including miR-21, miR-10b, miR-221, and miR-222, have shown significant diagnostic and prognostic potential. Integration of these biomarkers with multi-omics technologies and liquid biopsy approaches may improve early diagnosis, treatment monitoring, and prediction of tumor recurrence. However, standardized isolation methods and large multicenter validation studies remain essential before exosomal biomarkers can be routinely applied in clinical practice16.
3.4 Exosomal Biomarkers in Ischemic Stroke
Ischemic stroke is a leading cause of mortality and long-term disability worldwide, resulting from the interruption of cerebral blood flow and subsequent neuronal injury. Early diagnosis is critical because treatments such as thrombolysis and mechanical thrombectomy are highly time dependent. However, conventional imaging techniques provide limited information about the ongoing molecular events within the brain. Exosomes have emerged as promising liquid biopsy biomarkers because they are released rapidly by neurons, astrocytes, microglia, endothelial cells, and other brain cells after ischemic injury. Their ability to cross the blood brain barrier while carrying stable disease-specific cargo makes them valuable for early diagnosis, prognosis, and monitoring of therapeutic response17.
Following cerebral ischemia, both the concentration and molecular composition of circulating exosomes change significantly. Exosomal proteins such as neurofilament light chain (NfL), glial fibrillary acidic protein (GFAP), vascular endothelial growth factor (VEGF), brain-derived neurotrophic factor (BDNF), matrix metalloproteinase-9 (MMP-9), and inflammatory mediators reflect neuronal damage, blood–brain barrier disruption, angiogenesis, and tissue repair. In addition, exosomal microRNAs, including miR-21, miR-124, miR-126, miR-210, and miR-223, regulate apoptosis, neuroinflammation, angiogenesis, oxidative stress, and neuronal regeneration, making them attractive biomarkers for evaluating stroke severity and predicting functional recovery. Emerging evidence also highlights the diagnostic value of exosomal long non-coding RNAs and circular RNAs, while advances in proteomics and transcriptomics continue to identify novel molecular signatures associated with ischemic stroke.
Despite encouraging progress, several challenges remain before exosomal biomarkers can be implemented in routine clinical practice. Variations in exosome isolation methods, analytical techniques, and patient heterogeneity contribute to inconsistent findings across studies. Standardized protocols and large multicenter validation studies are therefore required to establish reliable biomarker panels. Nevertheless, exosomes represent a highly promising platform for minimally invasive diagnosis, prognostic assessment, and personalized management of ischemic stroke, with the potential to complement conventional neuroimaging and improve precision medicine approaches18.
3.5 Exosomal Biomarkers in Multiple Sclerosis
Multiple sclerosis (MS) is a chronic autoimmune disorder characterized by demyelination, neuroinflammation, and progressive neurological disability. Current diagnostic tools, including MRI and cerebrospinal fluid analysis, have limited ability to predict disease progression and treatment response. Exosomes have emerged as promising minimally invasive biomarkers because they carry disease-specific proteins and RNAs released from neurons, oligodendrocytes, astrocytes, microglia, and immune cells19.
Exosomes in MS contain elevated levels of neurofilament light chain (NfL), myelin basic protein (MBP), myelin oligodendrocyte glycoprotein (MOG), glial fibrillary acidic protein (GFAP), and inflammatory cytokines, reflecting axonal injury, demyelination, gliosis, and immune activation. Dysregulated exosomal miRNAs, particularly miR-155, miR-146a, and miR-219, are associated with neuroinflammation and remyelination, while lncRNAs and circRNAs further contribute to disease regulation. Multi-omics studies have also identified alterations in proteins, lipids, and metabolites linked to MS progression.
Circulating exosomes enable repeated, non-invasive monitoring of disease activity and therapeutic response through liquid biopsy. However, clinical application is limited by methodological variability, disease heterogeneity, and the lack of standardized isolation and analytical protocols. Overall, exosome-based biomarkers hold significant promise for improving the diagnosis, prognosis, and personalized management of multiple sclerosis, although further large-scale clinical validation is required20.
3.6 Exosomal Biomarkers in Amyotrophic Lateral Sclerosis
Amyotrophic lateral sclerosis (ALS) is a progressive neurodegenerative disease characterized by the loss of upper and lower motor neurons, resulting in muscle weakness, paralysis, and respiratory failure. Early diagnosis remains challenging because symptoms appear after significant neuronal loss. Exosomes have emerged as promising minimally invasive biomarkers as they carry disease-specific proteins and RNAs released from neurons, astrocytes, microglia, and muscle cells, reflecting ongoing neurodegeneration21.
Exosomal biomarkers in ALS include TDP-43, mutant SOD1, neurofilament light chain (NfL), neurofilament heavy chain (NfH), inflammatory cytokines, and heat shock proteins. Elevated exosomal TDP-43 and NfL are strongly associated with disease severity, motor neuron degeneration, and poor prognosis. Dysregulated exosomal miRNAs, including miR-206, miR-155, miR-124, miR-9, and miR-338-3p, regulate neuroinflammation, neuronal survival, and neuromuscular repair, while lncRNAs and circRNAs are emerging as additional biomarkers. Multi-omics studies have further identified disease-related alterations in proteins, lipids, metabolites, and RNAs involved in mitochondrial dysfunction, oxidative stress, and RNA metabolism.
Liquid biopsy using circulating exosomes enables repeated, non-invasive monitoring of disease progression and therapeutic response. However, clinical translation is limited by methodological variability, disease heterogeneity, and the lack of standardized analytical protocols. Overall, exosome-based biomarkers show strong potential for early diagnosis, prognosis, patient stratification, and personalized management of ALS, although large-scale clinical validation is still required22.
3.7 Exosomal Biomarkers in Huntington’s Disease
Huntington’s disease (HD) is an inherited neurodegenerative disorder caused by an expanded CAG repeat in the HTT gene, leading to the production of mutant huntingtin (mHTT) protein. Progressive accumulation of mHTT results in neuronal dysfunction, particularly within the striatum and cerebral cortex, causing motor impairment, cognitive decline, and psychiatric symptoms. Although genetic testing confirms the diagnosis, reliable biomarkers are still needed to monitor disease progression and therapeutic response.
Exosomes have emerged as promising liquid biopsy biomarkers because they can cross the blood–brain barrier and carry proteins, messenger RNAs, microRNAs (miRNAs), and other regulatory molecules that reflect the pathological state of neurons and glial cells.^154^ Neuron-derived exosomes containing mutant huntingtin protein, neurofilament light chain (NfL), brain-derived neurotrophic factor (BDNF), and inflammatory proteins have shown potential for assessing neuronal injury and disease severity. Elevated exosomal NfL correlates with neurodegeneration, while reduced BDNF is associated with impaired neuronal survival and cognitive decline.
Exosomal miRNAs, including miR-124, miR-9, miR-132, miR-196a, and miR-22, regulate pathways involved in neuronal differentiation, synaptic plasticity, apoptosis, mitochondrial function, and neuroinflammation. Changes in these miRNAs may help identify disease stage and monitor progression. In particular, miR-196a has attracted attention because of its potential role in reducing mutant huntingtin expression and limiting neurotoxicity.
Overall, exosome-based biomarkers provide a minimally invasive approach for monitoring Huntington’s disease by reflecting ongoing molecular changes associated with neurodegeneration. Continued advances in exosome isolation, multi-omics technologies, and standardized analytical methods are expected to improve their clinical utility for diagnosis, prognosis, and personalized therapeutic monitoring23.
3.8 Comparative Analysis of Exosomal Biomarkers: Proteomics, Transcriptomics, microRNAs, Long Non-Coding RNAs, Circular RNAs, Liquid Biopsy, Diagnostic Performance, Prognostic Significance, and Clinical Translation
Exosomes have emerged as powerful biomarker platforms for neurological disorders because they carry proteins, messenger RNAs, microRNAs (miRNAs), long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), lipids, and metabolites that reflect the physiological and pathological state of their parent cells. Their ability to cross the blood–brain barrier (BBB) and remain stable in peripheral blood makes them ideal candidates for minimally invasive liquid biopsy, enabling repeated monitoring of disease progression and therapeutic response.
Comparative studies across Alzheimer’s disease (AD), Parkinson’s disease (PD), glioblastoma (GBM), ischemic stroke (IS), multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), and Huntington’s disease (HD) demonstrate both shared and disease-specific exosomal biomarkers. Common proteins, including neurofilament light chain (NfL), glial fibrillary acidic protein (GFAP), heat shock proteins, inflammatory cytokines, and mitochondrial proteins, reflect neurodegeneration and neuroinflammation, whereas disease-specific biomarkers such as amyloid-β and phosphorylated tau (AD), α-synuclein (PD), EGFRvIII (GBM), myelin basic protein (MS), TDP-43 (ALS), and mutant huntingtin (HD) improve diagnostic specificity.
Advances in proteomics and transcriptomics have enabled comprehensive profiling of exosomal proteins and RNAs associated with neuronal injury, synaptic dysfunction, mitochondrial impairment, angiogenesis, immune regulation, and oxidative stress. Among RNA biomarkers, miRNAs remain the most extensively investigated because of their stability and ability to regulate multiple signaling pathways simultaneously. Likewise, lncRNAs and circRNAs provide additional information on gene regulation and disease-specific molecular mechanisms, and their combined analysis with proteins and miRNAs improves diagnostic accuracy.
Exosome-based liquid biopsy offers significant clinical advantages over conventional cerebrospinal fluid analysis and tissue biopsy. Exosomes isolated from plasma or serum provide real-time information on disease activity, prognosis, therapeutic response, and recurrence through a simple, minimally invasive blood sample. However, differences in exosome isolation techniques, analytical platforms, and biomarker normalization continue to limit reproducibility across studies. Therefore, standardized protocols, multicenter validation, and integration of multi-omics data with artificial intelligence are essential for successful clinical translation.
Overall, exosomal biomarkers represent a promising next-generation diagnostic platform for neurological disorders. Their unique molecular cargo, combined with advances in multi-omics technologies and computational analysis, supports their future application in early diagnosis, prognostic evaluation, disease monitoring, and precision medicine24.
4. ENGINEERING STRATEGIES FOR EXOSOMES
Exosomes have emerged as highly promising nanocarriers for targeted drug delivery because of their excellent biocompatibility, low immunogenicity, prolonged circulation time, natural ability to cross biological barriers, and efficient intercellular communication. Although native exosomes possess inherent therapeutic potential, their clinical application is often limited by low drug-loading efficiency, poor tissue specificity, heterogeneous cargo composition, rapid clearance, and variability related to donor cell sources. To address these challenges, a wide range of engineering strategies has been developed to improve cargo loading, targeting ability, biodistribution, and therapeutic efficacy.
Exosome engineering is broadly classified into pre-isolation (indirect) and post-isolation (direct) approaches. In pre-isolation engineering, donor cells are genetically or chemically modified before exosome secretion, allowing therapeutic molecules to be naturally incorporated into newly formed exosomes. In contrast, post-isolation engineering involves direct modification of purified exosomes using physical, chemical, or biological techniques to load therapeutic cargo or functionalize the exosomal surface. Each strategy offers distinct advantages depending on the therapeutic cargo, target tissue, and intended clinical application.
An ideal engineered exosome should exhibit high cargo-loading efficiency, prolonged circulation, low immunogenicity, efficient blood–brain barrier (BBB) penetration, selective targeting, controlled cargo release, and compatibility with large-scale Good Manufacturing Practice (GMP) production. Achieving these characteristics requires careful optimization of donor cell selection, loading methods, membrane modification, purification techniques, and quality control procedures.
Recent advances in genetic engineering, membrane functionalization, chemical conjugation, hybrid nanovesicle design, microfluidic technologies, and artificial intelligence-assisted optimization have significantly improved the development of engineered exosomes. These next-generation delivery systems have demonstrated considerable therapeutic potential in neurological disorders, including Alzheimer’s disease, Parkinson’s disease, glioblastoma, ischemic stroke, multiple sclerosis, amyotrophic lateral sclerosis, Huntington’s disease, spinal cord injury, and traumatic brain injury, particularly because of their ability to efficiently deliver therapeutic molecules across the BBB while reducing systemic toxicity.
Despite encouraging progress, several challenges remain before engineered exosomes can be widely adopted in clinical practice. Large-scale manufacturing, standardized engineering protocols, maintenance of membrane integrity, batch-to-batch consistency, long-term storage stability, regulatory approval, and cost-effective production continue to be important hurdles. Ongoing advances in bioengineering, nanotechnology, and pharmaceutical manufacturing are expected to overcome these limitations and accelerate the clinical translation of engineered exosome-based therapies for neurological diseases25,26.
4.1 Donor Cell Engineering
Donor cell engineering is a pre-isolation strategy in which parental cells are genetically modified before exosome production. This approach allows cells to naturally package therapeutic molecules such as miRNAs, siRNAs, mRNAs, proteins, and CRISPR/Cas components into exosomes while preserving their biological properties. It also enables the expression of targeting ligands (e.g., Lamp2b-RVG) to improve blood–brain barrier (BBB) penetration and brain-specific delivery.
Common donor cells include mesenchymal stem cells (MSCs), neural stem cells (NSCs), HEK293 cells, and immune cells. MSC-derived exosomes are most widely used because of their anti-inflammatory, neuroprotective, and regenerative properties. Although donor cell engineering provides high cargo-loading efficiency and precise targeting, challenges such as genetic stability, large-scale production, and batch-to-batch variability remain. Overall, it is a promising strategy for developing targeted exosome-based therapies for neurological disorders27.
4.2 Genetic Engineering of Exosomes
Genetic engineering is an advanced strategy that enhances the therapeutic potential of exosomes by precisely modifying their molecular cargo and membrane composition. Through recombinant DNA technology, gene editing, and RNA engineering, donor cells are engineered to produce exosomes enriched with therapeutic proteins, nucleic acids, and targeting ligands. This approach improves cargo loading, targeting specificity, and delivery efficiency, making genetically engineered exosomes highly promising for brain-targeted drug delivery.
Plasmid transfection and viral vectors, including lentiviral and adeno-associated viral (AAV) systems, are commonly used to introduce therapeutic genes into donor cells. These engineered cells subsequently secrete exosomes containing miRNAs, siRNAs, mRNAs, proteins, and other bioactive molecules with enhanced therapeutic efficacy. More recently, CRISPR/Cas-based genome editing has enabled exosome-mediated delivery of gene-editing components for correcting or silencing disease-associated genes involved in neurological disorders.
Genetic engineering also allows surface modification of exosomes by expressing fusion proteins such as Lamp2b or CD63 linked with targeting peptides including rabies virus glycoprotein (RVG), angiopep-2, and transferrin-binding peptides. These modifications significantly improve blood–brain barrier (BBB) penetration and selective uptake by neuronal cells. In addition, synthetic biology approaches using inducible promoters and programmable gene circuits enable controlled exosome production and stimulus-responsive therapeutic cargo release.
Despite its advantages, genetic engineering faces challenges including biosafety concerns associated with viral vectors, incomplete understanding of cargo-sorting mechanisms, manufacturing complexity, and batch-to-batch variability. Nevertheless, continuous advances in genome editing, RNA biology, synthetic biology, and artificial intelligence-assisted design are expected to improve the precision, scalability, and clinical translation of genetically engineered exosomes for neurological diseases28.
4.3 Surface Modification of Exosomes
Surface modification is an important engineering strategy that enhances the targeting ability, circulation time, and therapeutic efficacy of exosomes without significantly affecting their internal cargo. By modifying the exosomal membrane with targeting ligands, antibodies, peptides, polymers, or aptamers, engineered exosomes achieve improved biodistribution and efficient delivery across the blood–brain barrier (BBB).
Common surface modification methods include lipid insertion, click chemistry, PEGylation, antibody conjugation, and aptamer functionalization. Brain-targeting ligands such as rabies virus glycoprotein (RVG), angiopep-2, and transferrin-binding peptides significantly enhance neuronal uptake and BBB penetration. In addition, imaging probes and stimuli-responsive molecules can be incorporated for real-time tracking and controlled drug release.
Despite these advantages, challenges such as membrane instability, altered cellular uptake, manufacturing complexity, and large-scale standardization remain. Nevertheless, continued advances in nanotechnology, bioorthogonal chemistry, and synthetic biology are expected to improve the clinical translation of surface-engineered exosomes for neurological drug delivery29.
4.4 Chemical Conjugation
Chemical conjugation is a post-isolation strategy used to modify purified exosomes by attaching therapeutic molecules, targeting ligands, polymers, or imaging agents to their surface. Unlike genetic engineering, this method directly modifies isolated exosomes, allowing flexible and precise functionalization while preserving the encapsulated cargo. It improves targeting efficiency, blood–brain barrier (BBB) penetration, circulation time, and therapeutic performance in neurological disorders.
Common conjugation methods include EDC/NHS coupling, maleimide–thiol chemistry, click chemistry, and lipid insertion, which enable stable attachment of peptides, antibodies, aptamers, and drugs to the exosomal membrane. Polyethylene glycol (PEG) is also widely used to prolong circulation and reduce immune clearance. These modifications enhance cellular uptake and increase delivery specificity to diseased tissues.
Despite its advantages, chemical conjugation requires careful optimization because excessive modification may affect membrane integrity, stability, or biological activity. Standardized protocols and scalable manufacturing are essential for successful clinical translation30.
4.5 Ligand Functionalization
Ligand functionalization is an important strategy for improving the targeting ability of engineered exosomes. By attaching specific ligands to the exosomal surface, receptor-mediated recognition and cellular uptake are enhanced, leading to improved blood–brain barrier (BBB) penetration, selective accumulation in diseased tissues, and reduced off-target effects. This approach significantly increases the therapeutic potential of exosome-based drug delivery for neurological disorders.
Several ligands have been successfully used for brain targeting. Rabies virus glycoprotein (RVG) enhances neuronal uptake through nicotinic acetylcholine receptors, while Angiopep-2 promotes BBB transport by targeting low-density lipoprotein receptor-related protein-1 (LRP1). Transferrin, Apolipoprotein E (ApoE), and cyclic RGD (cRGD) peptides further improve brain and glioblastoma targeting through receptor-mediated endocytosis. In addition, antibodies, antibody fragments, and aptamers provide high targeting specificity and efficient therapeutic delivery.
Despite these advantages, ligand functionalization requires careful optimization because ligand density, receptor variability, immunogenicity, and manufacturing consistency can influence targeting efficiency. Continued advances in ligand design, synthetic biology, and scalable production are expected to accelerate the clinical translation of ligand-functionalized exosomes for precision neurological therapy31.
4.6 Membrane Hybridization
Membrane hybridization is an advanced exosome engineering strategy that combines the natural biological properties of exosomes with the advantages of synthetic nanocarriers. This approach improves drug-loading capacity, stability, targeting efficiency, and blood–brain barrier (BBB) penetration while maintaining the biocompatibility and low immunogenicity of native exosomes. As a result, hybrid vesicles have become promising carriers for brain-targeted drug delivery.
The most common approach is liposome–exosome fusion, which combines the high loading efficiency of liposomes with the natural targeting ability of exosomes. Hybridization can be achieved using freeze–thaw cycles, membrane extrusion, polyethylene glycol (PEG)-mediated fusion, or sonication. In addition, exosomes can be fused with membranes derived from red blood cells, macrophages, platelets, stem cells, or neuronal cells to enhance circulation time, tissue targeting, and therapeutic efficacy.
Despite these advantages, membrane hybridization requires careful optimization to preserve membrane integrity, achieve reproducible fusion, and ensure large-scale manufacturing. Continued advances in biomaterials, synthetic biology, and microfluidic technologies are expected to accelerate the clinical translation of membrane-hybridized exosomes for neurological disorders32.
4.7 Cargo Loading Methods
Cargo loading is a key step in exosome engineering because it determines the amount, stability, and therapeutic effectiveness of the delivered molecules. Native exosomes naturally carry proteins, lipids, messenger RNAs (mRNAs), microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs), but therapeutic applications often require additional cargo such as drugs, proteins, nucleic acids, or CRISPR/Cas components. Cargo loading methods are broadly classified into endogenous (pre-loading) and exogenous (post-loading) approaches. Endogenous loading modifies donor cells before exosome formation, resulting in efficient and stable cargo incorporation, whereas exogenous loading introduces therapeutic molecules into purified exosomes after isolation, offering greater flexibility for different cargo types33.
Common exogenous methods include passive incubation, electroporation, sonication, extrusion, freeze–thaw cycles, and microfluidic loading. Each technique has specific advantages and limitations depending on the cargo properties and required loading efficiency. An ideal loading method should maximize encapsulation while preserving exosome membrane integrity, biological activity, and blood–brain barrier (BBB) penetration.
Recent advances in microfluidics, biomaterials, and artificial intelligence-assisted optimization have improved loading efficiency, reproducibility, and scalability for clinical applications. Overall, selecting the appropriate cargo-loading strategy is essential for developing safe, effective, and clinically translatable engineered exosomes for neurological disorders34.
4.8 Electroporation
Electroporation is one of the most widely used active cargo-loading methods for engineered exosomes, particularly for loading small interfering RNA (siRNA), microRNA (miRNA), messenger RNA (mRNA), CRISPR/Cas components, proteins, and peptides. It uses short electrical pulses to create temporary pores in the exosomal membrane, allowing therapeutic molecules to enter before the membrane reseals. This approach is especially valuable for neurological disorders because it enables efficient delivery of nucleic acid therapeutics across the blood–brain barrier (BBB)35.
Electroporation provides higher loading efficiency than passive methods and has been successfully applied for delivering gene-silencing molecules targeting BACE1, SNCA, HTT, APP, SOD1, and other disease-associated genes. However, high electrical intensity may cause cargo aggregation, membrane damage, vesicle fusion, or reduced biological activity. Therefore, optimization of voltage, pulse duration, and buffer conditions is essential to maintain exosome integrity.
Recent advances such as microfluidic electroporation and continuous-flow systems have improved loading efficiency, reproducibility, and scalability while minimizing membrane disruption. Overall, electroporation remains one of the most reliable and clinically relevant techniques for engineering exosomes for brain-targeted drug delivery36.
4.9 Sonication
Sonication is an efficient active cargo-loading method that uses ultrasonic waves to temporarily increase the permeability of the exosomal membrane, allowing therapeutic molecules to enter before the membrane reseals. It is particularly effective for loading small-molecule drugs, proteins, peptides, microRNAs (miRNAs), messenger RNAs (mRNAs), and small interfering RNAs (siRNAs). Compared with passive loading, sonication generally provides higher encapsulation efficiency while maintaining the biological properties of exosomes.
This technique has been widely explored for neurological disorders, where sonicated exosomes have shown improved blood–brain barrier (BBB) penetration and enhanced delivery of therapeutic agents in Alzheimer's disease, Parkinson's disease, glioblastoma, and ischemic stroke models. However, excessive ultrasonic exposure may lead to membrane disruption, vesicle aggregation, or cargo leakage, making optimization of sonication conditions essential.
Recent advances in automated and microfluidic sonication systems have improved loading efficiency, reproducibility, and scalability for clinical translation. Overall, sonication is a reliable and versatile cargo-loading strategy for developing engineered exosomes for brain-targeted drug delivery37.
4.10 Extrusion
Extrusion is a widely used physical cargo-loading method that improves both therapeutic loading and exosome uniformity. In this technique, exosomes mixed with therapeutic cargo are repeatedly passed through nanoporous membranes under controlled pressure. This temporarily disrupts the phospholipid bilayer, allowing drugs or biomolecules to enter the vesicles before the membrane rapidly reassembles. As a result, extrusion provides high loading efficiency while producing exosomes with a uniform size distribution, which is beneficial for brain-targeted drug delivery.
Extrusion effectively loads both hydrophilic and hydrophobic drugs, as well as proteins, peptides, siRNAs, miRNAs, mRNAs, and CRISPR/Cas components. It also supports simultaneous loading of multiple therapeutics and is widely used to generate hybrid exosomes by fusing native exosomes with liposomes or other nanocarriers, enhancing cargo capacity, stability, and BBB penetration38.
Compared with passive incubation, extrusion generally offers higher loading efficiency and reduces nucleic acid aggregation compared with electroporation. However, repeated mechanical stress may alter membrane proteins or lipid organization, making careful optimization and post-loading characterization essential.
Recent advances in automated extrusion systems, microfluidics, and continuous-flow manufacturing have improved reproducibility and scalability, supporting Good Manufacturing Practice (GMP) production. Overall, extrusion is a versatile, scalable, and clinically promising strategy for engineering exosomes for neurological drug delivery39.
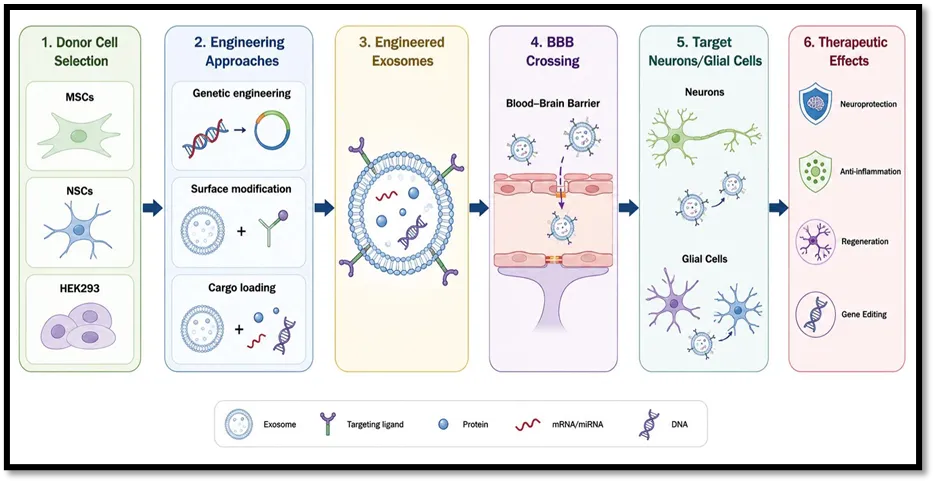
Fig No. 2 Engineering Strategies for Exosome-Based Brain Drug Delivery
4.11 Passive Incubation
Passive incubation is the simplest and most commonly used post-isolation cargo-loading method for engineered exosomes. In this approach, purified exosomes are incubated with therapeutic molecules under physiological conditions, allowing cargo to associate with or diffuse into exosomes through natural physicochemical interactions. Because no external physical or chemical force is applied, the structural integrity, membrane proteins, and biological properties of exosomes are largely preserved, making this method suitable for neurological drug delivery.
Passive incubation is particularly effective for loading hydrophobic small-molecule drugs such as curcumin, paclitaxel, doxorubicin, temozolomide, and rapamycin, which readily partition into the phospholipid bilayer. In contrast, hydrophilic drugs, proteins, peptides, siRNAs, mRNAs, and CRISPR/Cas components generally show lower loading efficiency and often require active loading methods. The technique is simple, cost-effective, scalable, and compatible with Good Manufacturing Practice (GMP) production while minimizing membrane damage, vesicle aggregation, and cargo degradation.
However, passive incubation has relatively low loading efficiency, particularly for large biomolecules, and prolonged incubation may be required to achieve adequate encapsulation. Recent improvements, including optimized temperature, pH, amphiphilic carriers, and microfluidic-assisted incubation, have enhanced cargo loading while preserving exosome integrity. Overall, passive incubation remains a valuable strategy for delivering hydrophobic therapeutics in brain-targeted exosome systems because of its simplicity, safety, and excellent preservation of native exosomal characteristics40.
4.12 Freeze–Thaw Method
The freeze–thaw method is a simple and widely used physical cargo-loading strategy that employs repeated freezing and thawing cycles to temporarily increase exosomal membrane permeability. During this process, transient membrane disruption allows therapeutic molecules to enter the exosome before the membrane reseals. Although its loading efficiency is generally lower than electroporation, sonication, or extrusion, the method is valued for its simplicity, low cost, and ability to preserve the biological properties of exosomes.
Freeze–thaw loading is compatible with a broad range of therapeutic cargos, including small-molecule drugs, proteins, peptides, nucleic acids, and nanoparticles. It is also widely used to generate hybrid exosomes by promoting membrane fusion between exosomes and liposomes, resulting in improved drug-loading capacity, enhanced BBB penetration, and better therapeutic performance in neurological disorders. Optimized protocols generally use three to six freeze–thaw cycles to balance loading efficiency with membrane integrity.
Despite these advantages, repeated freeze–thaw cycles may induce vesicle aggregation, membrane fusion, protein denaturation, and heterogeneous particle populations, making careful optimization essential. Recent advances in programmable cryogenic systems, membrane stabilizers, and combination approaches with extrusion or microfluidics have improved reproducibility and loading efficiency. Overall, the freeze–thaw method remains a practical and scalable strategy for exosome engineering, particularly for hybrid vesicle fabrication and brain-targeted drug delivery41.
4.13 Microfluidics-Based Engineering
Microfluidics-based engineering is an advanced exosome modification strategy that enables precise control over exosome isolation, cargo loading, membrane engineering, and large-scale production. By manipulating fluids within microscale channels under controlled conditions, microfluidic platforms improve loading efficiency, reduce batch-to-batch variability, minimize membrane damage, and enhance reproducibility compared with conventional methods such as electroporation, sonication, and extrusion.
Microfluidic systems support efficient loading of small-molecule drugs, proteins, peptides, siRNAs, miRNAs, mRNAs, CRISPR/Cas components, and imaging agents while preserving exosome integrity. They also facilitate membrane hybridization and surface functionalization, producing multifunctional exosomes with improved blood–brain barrier (BBB) penetration, targeting specificity, and therapeutic efficacy for neurological disorders.
An important advantage of microfluidics is the integration of multiple processes—including exosome isolation, purification, cargo loading, characterization, and quality control—within a single automated platform. Continuous-flow manufacturing improves scalability, sterility, and compliance with Good Manufacturing Practice (GMP) standards, making it highly suitable for clinical translation.
Recent advances integrating artificial intelligence (AI), machine learning, and real-time sensing have further optimized loading conditions, manufacturing consistency, and quality assurance. Although challenges such as device cost, fabrication complexity, and standardization remain, microfluidics represents one of the most promising technologies for developing next-generation engineered exosomes for brain-targeted drug delivery42.
5. BRAIN-TARGETED DRUG DELIVERY
5.1 Blood–Brain Barrier: Structure and Challenges
The blood–brain barrier (BBB) is a highly selective physiological barrier that protects the central nervous system (CNS) by regulating the movement of molecules between the bloodstream and brain tissue. While essential for maintaining brain homeostasis and preventing the entry of harmful substances, the BBB also limits the delivery of therapeutic agents. More than 98% of small-molecule drugs and nearly all biologics, including proteins, peptides, antibodies, and nucleic acids, cannot cross the intact BBB at therapeutic levels, making effective treatment of neurological disorders extremely challenging.
Structurally, the BBB consists of brain microvascular endothelial cells (BMECs) connected by tight junction proteins such as claudins, occludin, junctional adhesion molecules (JAMs), and zonula occludens (ZO) proteins. Together with pericytes, astrocytes, neurons, microglia, and the extracellular matrix, these cells form the neurovascular unit (NVU), which maintains BBB integrity and regulates nutrient transport, immune surveillance, and neuronal homeostasis. Specialized transporters, including GLUT1, LAT1, and transferrin receptors, facilitate the entry of essential nutrients, whereas efflux pumps such as P-glycoprotein (P-gp), breast cancer resistance protein (BCRP), and multidrug resistance-associated proteins (MRPs) actively remove many therapeutic agents, further limiting brain drug accumulation43.
The BBB poses a major obstacle in diseases such as Alzheimer’s disease, Parkinson’s disease, glioblastoma, multiple sclerosis, and ischemic stroke, where many promising therapeutics fail because of poor brain penetration. Although approaches such as intracerebral injection, focused ultrasound, osmotic BBB disruption, and synthetic nanoparticles have been explored, they are often limited by invasiveness, toxicity, or poor targeting efficiency.
Engineered exosomes have emerged as promising brain-targeted nanocarriers because of their small size, excellent biocompatibility, low immunogenicity, and natural ability to cross the BBB through receptor-mediated transport. Surface modification with targeting ligands such as RVG, Angiopep-2, transferrin, and ApoE further enhances BBB penetration and selective delivery to diseased brain tissues. Ongoing advances in exosome engineering, cargo loading, and membrane functionalization continue to improve their potential for treating neurological disorders44.
5.2 Mechanisms of Blood–Brain Barrier Transport
Transport across the blood–brain barrier (BBB) is essential for delivering therapeutics to the central nervous system (CNS). The BBB tightly regulates molecular exchange through specialized transport pathways that maintain brain homeostasis while preventing the entry of harmful substances. Engineered exosomes cross the BBB by utilizing these natural transport mechanisms rather than disrupting barrier integrity, making them highly promising carriers for brain-targeted drug delivery.
Paracellular diffusion contributes minimally to drug transport because brain endothelial cells are connected by tight junction proteins, including claudins, occludin, junctional adhesion molecules (JAMs), and zonula occludens (ZO) proteins. Although BBB permeability may increase in conditions such as ischemic stroke, neuroinflammation, and glioblastoma, relying on this pathway is unreliable and may increase the risk of neurotoxicity.
Transcellular passive diffusion allows only small lipophilic molecules to cross endothelial cells, whereas most therapeutic proteins, peptides, nucleic acids, and nanoparticles cannot penetrate efficiently. In addition, efflux transporters such as P-glycoprotein (P-gp), breast cancer resistance protein (BCRP), and multidrug resistance-associated proteins (MRPs) actively remove many drugs from the brain, further limiting therapeutic efficacy.
Among all transport pathways, receptor-mediated transcytosis (RMT) is the primary mechanism exploited by engineered exosomes. By functionalizing exosomal membranes with ligands such as rabies virus glycoprotein (RVG), Angiopep-2, transferrin, and apolipoprotein E (ApoE), exosomes bind to receptors on brain endothelial cells, undergo endocytosis, and are transported across the BBB into brain tissue. This pathway provides efficient and selective delivery while maintaining BBB integrity.
Adsorptive-mediated transcytosis (AMT) is another pathway that depends on electrostatic interactions between positively charged molecules and the negatively charged endothelial surface. Although less specific than RMT, AMT can enhance exosome uptake when surface charge is carefully optimized45.
Following endothelial binding, exosomes are internalized through clathrin-mediated endocytosis, caveolin-dependent endocytosis, macropinocytosis, or lipid raft-mediated uptake. Their membrane proteins and lipids, including CD9, CD63, CD81, integrins, sphingomyelin, cholesterol, and phosphatidylserine, influence cellular recognition, intracellular trafficking, and successful transport into the brain.
Recent advances in microfluidic BBB models, organ-on-a-chip systems, live-cell imaging, and artificial intelligence (AI)-based computational modeling have improved understanding of exosome transport mechanisms and are helping optimize next-generation brain-targeted delivery systems46.
5.3 Rabies Virus Glycoprotein (RVG) Peptide-Mediated Brain Targeting
Rabies virus glycoprotein (RVG) peptide is one of the most extensively investigated targeting ligands for engineering exosomes capable of crossing the blood–brain barrier (BBB). Derived from the neurotropic rabies virus glycoprotein, RVG retains strong neuronal targeting ability without viral infectivity. Surface modification of exosomes with RVG significantly enhances BBB penetration and selective delivery of therapeutic cargo to neurons, astrocytes, and other central nervous system (CNS) cells, making RVG-functionalized exosomes a promising platform for neurological drug delivery.
RVG primarily binds to nicotinic acetylcholine receptors (nAChRs) expressed on brain microvascular endothelial cells and neurons. This interaction promotes receptor-mediated transcytosis across the BBB, followed by efficient neuronal uptake after entry into the brain. Consequently, RVG enhances both BBB transport and neuron-specific targeting, providing greater therapeutic specificity than many conventional targeting ligands.
RVG peptides are commonly displayed on exosomes through genetic engineering of donor cells by fusing RVG with exosomal membrane proteins such as Lamp2b, CD63, or CD9. Alternatively, post-isolation chemical conjugation methods, including click chemistry, PEG-mediated coupling, and lipid insertion, enable controlled surface functionalization while preserving exosome morphology. Both strategies have demonstrated effective brain-targeted delivery of therapeutic cargo.
RVG-engineered exosomes have successfully delivered siRNAs, miRNAs, mRNAs, CRISPR/Cas components, proteins, enzymes, and small-molecule drugs in numerous preclinical studies. In Alzheimer's disease, RVG-modified exosomes improve delivery of anti-amyloid therapeutics and neuroprotective molecules, reducing amyloid deposition, neuroinflammation, and cognitive impairment. In Parkinson's disease, they efficiently transport α-synuclein-targeting siRNAs, neurotrophic factors, and antioxidant agents, protecting dopaminergic neurons and improving motor function. Similar therapeutic benefits have been reported in glioblastoma, ischemic stroke, traumatic brain injury, spinal cord injury, and amyotrophic lateral sclerosis.
Despite these advantages, RVG-mediated targeting has several limitations. Receptor saturation following repeated administration, expression of nAChRs in peripheral tissues, variability in ligand density, and manufacturing challenges may influence targeting efficiency and biodistribution. Therefore, standardized methods for ligand conjugation, exosome characterization, and large-scale production remain essential for clinical translation.
Recent advances include multifunctional exosomes incorporating RVG together with other BBB-targeting ligands such as Angiopep-2, transferrin, or ApoE peptides, as well as integration with imaging probes and stimuli-responsive delivery systems. These strategies further improve BBB penetration, targeting specificity, and therapeutic efficacy.
Overall, RVG peptide-mediated targeting remains one of the most effective approaches for brain-targeted exosome delivery. Its ability to promote receptor-mediated transcytosis and neuron-selective uptake has established RVG-functionalized exosomes as a leading platform for delivering nucleic acids, proteins, and therapeutic drugs to the CNS, supporting their continued development for neurological disorders47.
5.4 Angiopep-2-Mediated Brain Targeting
Angiopep-2 is one of the most effective targeting peptides used to enhance blood–brain barrier (BBB) penetration of engineered exosomes. It is a synthetic 19-amino-acid peptide derived from the Kunitz protease inhibitor domain of aprotinin and specifically targets the low-density lipoprotein receptor-related protein-1 (LRP1), which is highly expressed on brain microvascular endothelial cells, neurons, astrocytes, and glioma cells. By exploiting LRP1-mediated receptor-dependent transcytosis, Angiopep-2-functionalized exosomes efficiently cross the BBB and subsequently accumulate within neural tissues without disrupting BBB integrity.
Angiopep-2 is commonly displayed on exosomal membranes through genetic engineering of donor cells using membrane proteins such as Lamp2b or CD63, or by post-isolation chemical conjugation methods including click chemistry, lipid insertion, and PEG-mediated coupling. These approaches improve BBB targeting while preserving exosome stability and biological activity.
Angiopep-2-engineered exosomes have shown significant therapeutic potential in glioblastoma, Alzheimer’s disease, Parkinson’s disease, and ischemic stroke. They efficiently deliver chemotherapeutic drugs, proteins, siRNAs, miRNAs, mRNAs, CRISPR/Cas systems, and neuroprotective molecules, resulting in enhanced BBB penetration, increased neuronal or tumor-cell uptake, improved therapeutic efficacy, and reduced systemic toxicity. In glioblastoma, Angiopep-2 provides dual targeting by facilitating BBB transport and selective accumulation in LRP1-overexpressing tumor cells.
Compared with many other targeting ligands, Angiopep-2 offers high transport efficiency through the abundant LRP1 pathway. However, because LRP1 is also expressed in peripheral tissues, some off-target uptake may occur, making optimization of ligand density and dosing important. Recent advances combining Angiopep-2 with RVG, transferrin, ApoE, imaging probes, and stimuli-responsive systems have further enhanced targeting specificity and clinical potential. Overall, Angiopep-2-mediated engineering represents a highly promising strategy for brain-targeted exosome delivery and future neurological therapeutics48.
5.5 Transferrin-Mediated Brain Targeting
Transferrin-mediated targeting is one of the most established strategies for enhancing blood–brain barrier (BBB) transport of engineered exosomes. This approach exploits transferrin receptor 1 (TfR1/CD71), which is highly expressed on brain microvascular endothelial cells (BMECs) and regulates iron transport into the central nervous system (CNS). By utilizing TfR1-mediated receptor-mediated transcytosis, transferrin-functionalized exosomes efficiently cross the BBB while maintaining its structural integrity, making them promising carriers for brain-targeted drug delivery.
Transferrin is incorporated onto exosomal membranes through genetic engineering of donor cells using membrane proteins such as Lamp2b or CD63, or by chemical conjugation methods including click chemistry, PEG-mediated coupling, lipid insertion, and carbodiimide chemistry. These approaches improve BBB targeting while preserving exosome stability and biological activity.
Transferrin-engineered exosomes have demonstrated significant therapeutic potential in glioblastoma, Alzheimer's disease, Parkinson's disease, and ischemic stroke. They effectively deliver chemotherapeutic drugs, proteins, siRNAs, miRNAs, mRNAs, CRISPR/Cas systems, and neuroprotective molecules, resulting in enhanced BBB penetration, increased neuronal or tumor-cell uptake, improved therapeutic efficacy, and reduced systemic toxicity. In glioblastoma, transferrin targeting enhances selective accumulation in transferrin receptor-overexpressing tumor cells.
Compared with passive targeting strategies, transferrin-mediated delivery provides efficient receptor-mediated transport of large biomolecules across the BBB. However, competition with endogenous transferrin, receptor saturation, and TfR1 expression in peripheral tissues may reduce brain specificity, highlighting the importance of optimizing ligand density and dosing. Recent advances combining transferrin with RVG, Angiopep-2, ApoE, imaging probes, and stimuli-responsive systems have further improved targeting efficiency and translational potential. Overall, transferrin-mediated engineering remains a clinically relevant and promising strategy for exosome-based brain-targeted drug delivery49.
5.6 Apolipoprotein E (ApoE)-Mediated Brain Targeting
Apolipoprotein E (ApoE)-mediated targeting enhances blood–brain barrier (BBB) transport of engineered exosomes by exploiting LDLR, LRP1, ApoER2, and VLDLR expressed on brain microvascular endothelial cells and neural cells. ApoE peptides are incorporated onto exosomes through genetic engineering or chemical conjugation, enabling efficient receptor-mediated transcytosis and targeted delivery to the brain.
ApoE-functionalized exosomes have shown therapeutic potential in Alzheimer's disease, Parkinson's disease, glioblastoma, ischemic stroke, and spinal cord injury by delivering drugs, proteins, siRNAs, miRNAs, and CRISPR/Cas systems, resulting in improved BBB penetration, enhanced neuronal uptake, reduced neuroinflammation, and increased therapeutic efficacy. Although peripheral receptor expression may cause off-target uptake, multifunctional systems combining ApoE with RVG, Angiopep-2, or transferrin further improve brain targeting. Overall, ApoE-mediated engineering is a promising strategy for exosome-based brain-targeted drug delivery50.
5.8 Intranasal Drug Delivery
Intranasal drug delivery is a promising non-invasive strategy for delivering engineered exosomes to the central nervous system (CNS) by bypassing the blood–brain barrier (BBB). Unlike intravenous administration, intranasal delivery enables direct nose-to-brain transport through the olfactory and trigeminal nerve pathways, reducing peripheral distribution and first-pass metabolism. The nanoscale size, biocompatibility, low immunogenicity, and cargo-protective properties of exosomes make them highly suitable carriers for this route.
Following intranasal administration, exosomes penetrate the nasal mucosa through transcellular transport, endocytosis, and extracellular pathways before reaching the brain via olfactory and trigeminal nerves. This allows efficient delivery of therapeutic proteins, nucleic acids, peptides, and small-molecule drugs to various brain regions without disrupting BBB integrity.
Preclinical studies have demonstrated that intranasally administered engineered exosomes improve therapeutic outcomes in Alzheimer's disease, Parkinson's disease, ischemic stroke, glioblastoma, and traumatic brain injury by reducing neuroinflammation, promoting neuronal survival, enhancing neuroregeneration, and improving functional recovery. Compared with systemic administration, this approach increases brain accumulation while minimizing off-target toxicity.
Despite these advantages, challenges such as mucociliary clearance, limited residence time, enzymatic degradation, and interindividual variability in nasal anatomy may reduce delivery efficiency. Strategies including mucoadhesive polymers, hydrogels, and surface modification with targeting ligands such as RVG or lactoferrin have been developed to enhance nose-to-brain transport.
Overall, intranasal administration represents a safe, patient-friendly, and effective route for brain-targeted delivery of engineered exosomes. Continued optimization of formulation design, pharmacokinetics, and large-scale clinical evaluation is expected to facilitate its translation for the treatment of neurological disorders51.
5.9 Intravenous Administration
Intravenous (IV) administration is the most widely investigated route for delivering engineered exosomes to the central nervous system (CNS). It enables controlled systemic delivery but requires exosomes to survive circulation, evade mononuclear phagocyte system (MPS) clearance, cross the blood–brain barrier (BBB), and accumulate in target brain tissues. Their nanoscale size, phospholipid bilayer, and low immunogenicity protect therapeutic cargo and support prolonged circulation.
Brain delivery primarily occurs through receptor-mediated transcytosis, which is enhanced by surface ligands such as RVG, Angiopep-2, transferrin, ApoE, lactoferrin, and T7 peptide. Surface engineering approaches including PEGylation, CD47 modification, and multifunctional ligand decoration further improve circulation time, reduce immune clearance, and enhance BBB penetration.
Intravenously administered engineered exosomes have shown therapeutic efficacy in Alzheimer’s disease, Parkinson’s disease, glioblastoma, ischemic stroke, traumatic brain injury, multiple sclerosis, and amyotrophic lateral sclerosis by delivering proteins, nucleic acids, neurotrophic factors, and small-molecule drugs, resulting in reduced neuroinflammation, enhanced neuroprotection, and improved functional recovery.
However, rapid uptake by the liver, spleen, and other peripheral organs, protein corona formation, receptor variability, and off-target distribution remain major limitations. Advances in biomimetic surface engineering, multifunctional targeting, AI-assisted optimization, and scalable Good Manufacturing Practice (GMP)-compliant production are expected to improve pharmacokinetics and accelerate clinical translation.
Overall, intravenous administration remains the principal route for engineered exosome-based brain drug delivery because of its clinical feasibility, precise dose control, and compatibility with targeted exosome engineering strategies for neurological disorders52.,
6. THERAPEUTIC APPLICATIONS OF ENGINEERED EXOSOMES IN NEUROLOGICAL DISORDERS
Engineered exosomes have emerged as promising nanotherapeutic platforms for neurological disorders because of their excellent biocompatibility, low immunogenicity, prolonged circulation, and ability to cross the blood–brain barrier (BBB). They can efficiently deliver diverse therapeutic cargo, including proteins, peptides, messenger RNA (mRNA), microRNA (miRNA), small interfering RNA (siRNA), long non-coding RNA (lncRNA), CRISPR/Cas gene-editing systems, enzymes, and small-molecule drugs, enabling targeted treatment of central nervous system (CNS) diseases.
Beyond drug delivery, engineered exosomes possess intrinsic biological activities that regulate neuroinflammation, oxidative stress, apoptosis, angiogenesis, synaptic plasticity, axonal regeneration, and immune responses. Their therapeutic performance can be further enhanced through cargo engineering, surface functionalization, receptor-specific targeting, and stimuli-responsive release. Exosomes derived from mesenchymal stem cells, neural stem cells, dendritic cells, macrophages, and induced pluripotent stem cells exhibit distinct therapeutic properties that support disease-specific applications.
Preclinical studies have demonstrated promising outcomes in neurodegenerative diseases, brain tumors, cerebrovascular disorders, and traumatic neurological injuries. However, therapeutic efficacy depends on factors such as donor-cell source, cargo selection, targeting ligands, administration route, and disease pathology. Continued optimization of these parameters and large-scale clinical validation will be essential for the successful translation of engineered exosome-based therapies into neurological practice.
6.1 Alzheimer’s Disease
Alzheimer’s disease (AD) is the most common neurodegenerative disorder and is characterized by progressive cognitive decline, amyloid-β (Aβ) plaque deposition, tau hyperphosphorylation, neuroinflammation, oxidative stress, mitochondrial dysfunction, synaptic loss, and neuronal degeneration. The limited efficacy of current therapies and poor blood–brain barrier (BBB) permeability have highlighted engineered exosomes as promising therapeutic carriers.
Engineered exosomes have been widely investigated for delivering small interfering RNA (siRNA), microRNA (miRNA), proteins, enzymes, and CRISPR/Cas gene-editing systems targeting key pathological pathways. Exosome-mediated delivery of BACE1 siRNA reduces amyloid-β production, while therapeutic miRNAs regulate neuroinflammation, apoptosis, oxidative stress, and synaptic function. Similarly, exosomes carrying anti-tau nucleic acids decrease tau aggregation and neuronal damage.
Engineered exosomes have also been used to deliver neprilysin, insulin-degrading enzyme, brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), glial cell line-derived neurotrophic factor (GDNF), and antioxidant enzymes, resulting in enhanced amyloid clearance, neuroprotection, improved mitochondrial function, and cognitive recovery in preclinical models. Surface modification with RVG peptide, Angiopep-2, transferrin, ApoE-derived peptides, and lactoferrin further improves BBB penetration and neuron-specific delivery.
Both intravenous and intranasal administration have demonstrated effective brain delivery, reduced amyloid and tau pathology, attenuation of neuroinflammation, and improved cognitive performance. However, challenges including manufacturing standardization, large-scale production, cargo-loading efficiency, long-term safety, and clinical validation remain to be addressed.
Overall, engineered exosomes represent a promising multifunctional therapeutic platform for Alzheimer’s disease by simultaneously targeting multiple pathogenic mechanisms and enabling efficient brain-specific delivery of diverse therapeutic cargos53.
6.2 Parkinson’s Disease
Parkinson's disease is characterized by progressive loss of dopaminergic neurons, α-synuclein aggregation, neuroinflammation, oxidative stress, and mitochondrial dysfunction. Current treatments improve symptoms but do not stop disease progression. Engineered exosomes can cross the blood–brain barrier (BBB) and deliver therapeutic cargo directly to affected brain regions.
Engineered exosomes carrying siRNAs, microRNAs, neurotrophic factors (GDNF, BDNF), antioxidant enzymes, anti-inflammatory molecules, and CRISPR/Cas systems reduce α-synuclein aggregation, protect dopaminergic neurons, suppress neuroinflammation, improve mitochondrial function, and restore motor function. Surface modification with RVG, Angiopep-2, transferrin, or ApoE enhances BBB penetration and neuron-specific targeting. Both intravenous and intranasal administration have shown promising results in preclinical studies.
Despite encouraging findings, challenges such as large-scale manufacturing, standardization, and long-term safety remain. Overall, engineered exosomes represent a promising disease-modifying strategy for Parkinson’s disease by enabling targeted and efficient brain delivery of therapeutic agents54.
6.3 Glioblastoma
Glioblastoma (GBM) is the most aggressive primary brain tumor with poor prognosis due to rapid growth, tumor heterogeneity, therapeutic resistance, and limited drug penetration across the blood–brain barrier (BBB). Engineered exosomes have emerged as promising nanocarriers because they efficiently cross the BBB and selectively deliver therapeutic agents to glioblastoma cells.
Engineered exosomes loaded with chemotherapeutic drugs, siRNAs, microRNAs, mRNAs, and CRISPR/Cas systems inhibit tumor growth, induce apoptosis, suppress angiogenesis, and overcome drug resistance. Surface modification with Angiopep-2, cRGD, transferrin, RVG, and ApoE enhances BBB penetration and tumor-specific targeting. Exosomes also modulate the tumor microenvironment by reducing immunosuppression and improving antitumor immune responses.
Both intravenous and intranasal administration have shown effective tumor targeting in preclinical studies. However, challenges including tumor heterogeneity, large-scale manufacturing, standardization, and long-term clinical safety remain. Overall, engineered exosomes represent a promising targeted therapeutic platform for glioblastoma by combining efficient brain delivery with multifunctional anticancer activity55.
6.4 Ischemic Stroke
Ischemic stroke occurs due to blockage of cerebral blood vessels, causing neuronal death through oxidative stress, inflammation, apoptosis, and blood–brain barrier (BBB) disruption. Engineered exosomes have emerged as promising therapeutics because they efficiently cross the BBB and deliver bioactive molecules such as miRNAs, siRNAs, proteins, antioxidants, and growth factors directly to ischemic brain tissue. These exosomes reduce neuroinflammation, protect neurons, restore BBB integrity, promote angiogenesis, stimulate neurogenesis, and enhance functional recovery. Surface modification with targeting ligands (e.g., RVG and Angiopep-2) further improves brain-specific delivery. Although preclinical studies have shown significant neuroprotective and regenerative effects, challenges including large-scale manufacturing, standardization, long-term safety, and clinical validation must be addressed before routine clinical application56.
6.5 Multiple Sclerosis
Multiple sclerosis (MS) is a chronic autoimmune disease characterized by inflammation, demyelination, and neurodegeneration in the CNS. Engineered exosomes show great therapeutic potential by crossing the blood–brain barrier (BBB) and delivering anti-inflammatory cytokines, miRNAs, growth factors, and gene-editing molecules. They suppress autoreactive immune responses, promote regulatory T cells, reduce neuroinflammation, enhance remyelination by stimulating oligodendrocyte precursor cells, and protect neurons from oxidative stress and apoptosis. Surface-modified exosomes improve targeted brain delivery, while both intravenous and intranasal routes have shown promising results. Although preclinical studies are encouraging, challenges such as large-scale production, standardization, safety, and clinical validation remain before routine clinical use57.
7. CLINICAL TRANSLATION OF ENGINEERED EXOSOMES
Engineered exosomes have shown significant therapeutic potential in neurological disorders due to their ability to cross the blood–brain barrier (BBB), deliver diverse therapeutic cargos, and exhibit low immunogenicity. However, clinical translation remains challenging because of variability in donor cells, cargo loading, manufacturing, purification, storage, and quality control. Standardized Good Manufacturing Practice (GMP) production, scalable manufacturing, and regulatory approval are essential for successful clinical application. Ongoing advances in engineering, quality assessment, and clinical research are expected to accelerate the development of safe and effective exosome-based therapies for neurological diseases58.
7.1 Current Clinical Trials
Most current clinical trials are Phase I or Phase I/II studies evaluating the safety, feasibility, dosing, and preliminary efficacy of mesenchymal stem cell-derived and engineered exosomes for neurological disorders such as ischemic stroke, Alzheimer’s disease, Parkinson’s disease, spinal cord injury, and amyotrophic lateral sclerosis. Early studies indicate good safety and tolerability, while engineered exosomes with targeted ligands and therapeutic cargos are showing improved therapeutic potential. However, larger randomized clinical trials and standardized protocols are still required to confirm long-term safety and clinical efficacy59.
7.2 Good Manufacturing Practice (GMP) Production
Good Manufacturing Practice (GMP) ensures that engineered exosomes are produced safely, consistently, and with high quality for clinical use. Standardized donor-cell selection, controlled culture conditions, reliable cargo loading, purification, and quality testing are essential to produce reproducible and contamination-free exosomes. Although GMP-compliant manufacturing has improved significantly, large-scale, cost-effective production remains a major challenge for the clinical translation of engineered exosome therapies60.
7.3 Large-Scale Production and Scalability
Large-scale production of engineered exosomes is essential for clinical application but remains technically challenging. Advanced bioreactors, three-dimensional cell culture, microfluidic cargo loading, and scalable purification methods improve exosome yield, quality, and consistency. Future automation and artificial intelligence-assisted manufacturing are expected to reduce costs and enable commercial-scale production61.
7.4 Isolation, Purification, and Quality Control
Isolation and purification are essential to obtain pure, functional, and safe engineered exosomes. Common methods include differential ultracentrifugation, size-exclusion chromatography (SEC), and tangential flow filtration (TFF), with TFF being most suitable for large-scale GMP production. Quality control involves assessing particle size, purity, sterility, morphology, cargo, and biological activity according to MISEV guidelines to ensure consistent clinical-grade exosomes62.
7.5 Storage, Stability, and Formulation
Proper storage is essential to maintain the stability and therapeutic activity of engineered exosomes. Storage at −80°C, minimizing freeze–thaw cycles, and using cryoprotectants such as trehalose improve exosome stability. Lyophilization (freeze-drying) and optimized formulations further enhance shelf life, transportation, and clinical applicability63.
7.6 Regulatory Considerations
Regulatory approval of engineered exosomes requires strict evaluation of their quality, safety, efficacy, and manufacturing consistency. Agencies such as the FDA and EMA regulate exosome therapies under biological or advanced therapy medicinal product (ATMP) frameworks. Standardized characterization, GMP production, preclinical safety, and well-designed clinical trials are essential for regulatory approval60.
7.7 Commercialization and Industrial Landscape
The commercialization of engineered exosomes is rapidly expanding, driven by advances in biotechnology, scalable manufacturing, and targeted drug delivery. Biotechnology companies are developing GMP-compliant production, purification, and engineering platforms, while growing investment and collaborations are accelerating clinical translation. However, high manufacturing costs, regulatory requirements, and reimbursement challenges remain major barriers to widespread clinical adoption64.
7.8 Current Challenges and Barriers to Clinical Translation
Despite significant advances in engineered exosome research, several biological, technological, manufacturing, regulatory, clinical, and economic challenges continue to hinder their clinical translation. Addressing these barriers is essential for achieving standardized, safe, and effective exosome-based therapies for neurological disorders.
A major challenge is the biological heterogeneity of exosomes, as their composition varies with donor-cell source, culture conditions, isolation methods, and engineering strategies, resulting in batch-to-batch variability and inconsistent therapeutic potency. Similarly, cargo-loading efficiency remains suboptimal because current techniques, including electroporation, sonication, extrusion, and microfluidics, differ in loading capacity and may compromise membrane integrity or endogenous cargo.
Another limitation is biodistribution and pharmacokinetics. Following systemic administration, many exosomes accumulate in the liver, spleen, and lungs rather than the brain. Although targeting ligands such as RVG, Angiopep-2, transferrin, ApoE, and lactoferrin improve brain delivery, targeting efficiency, clearance mechanisms, and long-term tissue distribution remain incompletely understood. Long-term safety also requires further investigation, particularly regarding repeated administration, off-target effects, immunogenicity, and the delivery of gene-editing cargo.
Manufacturing standardization remains a major obstacle because reproducible large-scale GMP production, purification, characterization, and storage protocols have not yet been fully established. In parallel, regulatory uncertainty persists due to the absence of exosome-specific guidelines for product classification, potency testing, quality control, and long-term safety assessment. Harmonized international regulatory frameworks are therefore needed.
Clinical evidence is still limited, with most studies remaining in early-phase trials involving small patient populations and short follow-up periods. Large, randomized multicenter studies are required to establish optimal dosing, administration routes, treatment schedules, biomarkers, and clinical endpoints. Furthermore, economic and logistical challenges, including high manufacturing costs, quality-control requirements, cold-chain storage, and limited shelf life, restrict large-scale commercialization.
Future progress will depend on integrating artificial intelligence, machine learning, multi-omics, computational modeling, biomaterials science, and precision medicine to optimize donor-cell selection, cargo loading, targeting, and patient-specific therapeutic design. Overall, overcoming biological variability, manufacturing and regulatory barriers, safety concerns, and economic limitations will be crucial for translating engineered exosomes into widely accessible therapies for neurological diseases59.
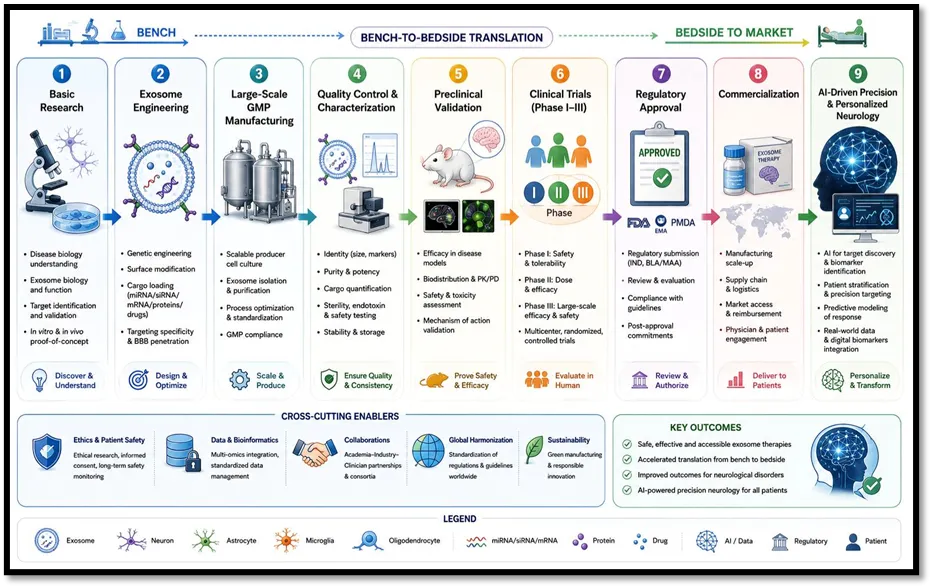
Fig No. 3 Future Roadmap for Clinical Translation of Engineered Exosomes
8. RESEARCH GAP
Despite significant advances in engineered exosome research, several key scientific and translational gaps continue to limit their clinical application in neurological disorders. A major challenge is the lack of standardized protocols for exosome production, isolation, purification, cargo loading, characterization, and therapeutic evaluation, resulting in poor reproducibility across studies despite the availability of MISEV guidelines.
Another important gap is cargo-loading efficiency and stability. Current loading techniques, including electroporation, sonication, extrusion, passive incubation, and microfluidics, show variable efficiency and may compromise membrane integrity or cargo retention. In addition, biodistribution, tissue targeting, pharmacokinetics, and pharmacodynamics remain incompletely understood, as many exosomes accumulate in off-target organs and their circulation time, cellular uptake, clearance, and dose–response relationships are not fully characterized.
Long-term safety and immunogenicity also require further investigation, particularly for repeated administration and genetically engineered exosomes carrying therapeutic nucleic acids or gene-editing systems. The influence of donor-cell heterogeneity on exosome composition and therapeutic efficacy remains insufficiently explored, while manufacturing variability and the absence of universally accepted potency and quality standards continue to hinder large-scale GMP production.
Further challenges include regulatory uncertainty, limited randomized clinical trials, and insufficient evidence regarding long-term efficacy and safety. In addition, cost-effectiveness, healthcare implementation, and commercialization remain poorly evaluated because of high manufacturing costs and complex production processes. Future studies should integrate artificial intelligence, multi-omics technologies, computational modeling, and precision medicine to optimize exosome engineering, targeting, and manufacturing. Greater interdisciplinary collaboration and international standardization will be essential to bridge these research gaps and accelerate the safe clinical translation of engineered exosomes for neurological disorders.
9. DISCUSSION
Engineered exosomes have emerged as promising nanocarriers for neurological disorders because of their excellent biocompatibility, low immunogenicity, intrinsic ability to cross the blood–brain barrier (BBB), and capacity to deliver diverse therapeutic cargos, including proteins, RNAs, and small-molecule drugs. Their natural origin and multifunctional properties make them superior to many conventional drug delivery systems.
Recent advances in donor-cell engineering, surface functionalization, and cargo-loading techniques have significantly improved targeting efficiency and therapeutic performance. Engineered exosomes can simultaneously regulate inflammation, oxidative stress, apoptosis, neuroregeneration, and synaptic repair, making them suitable for complex neurological diseases such as Alzheimer's disease, Parkinson's disease, stroke, and glioblastoma. In addition, exosome-derived biomarkers have shown considerable potential for early diagnosis and disease monitoring through minimally invasive liquid biopsy.
Despite these advances, several limitations remain. Variability in donor-cell sources, isolation methods, engineering strategies, and dosing protocols results in inconsistent therapeutic outcomes and limits comparison among studies. Large-scale GMP manufacturing, standardized quality control, efficient cargo loading, and long-term storage also remain major technical challenges. Furthermore, limited knowledge of pharmacokinetics, biodistribution, repeated-dose safety, and long-term immunogenicity continues to hinder clinical translation.
Current clinical evidence is still limited, with most studies remaining in early-phase clinical trials. Future research should prioritize standardized manufacturing protocols, robust clinical validation, improved brain-targeting strategies, and harmonized regulatory guidelines. Integration of artificial intelligence, multi-omics technologies, and advanced bioengineering may further optimize exosome design, manufacturing, and personalized therapeutic applications.
Overall, engineered exosomes represent a highly promising platform for brain-targeted drug delivery and precision neurology. However, overcoming challenges related to standardization, manufacturing scalability, regulatory approval, and long-term safety will be essential before these therapies can become part of routine clinical practice.
10. FUTURE PERSPECTIVE
Engineered exosomes are expected to play a pivotal role in the future of precision medicine for neurological disorders. Advances in molecular biology, nanotechnology, synthetic biology, and regenerative medicine are likely to transform engineered exosomes into standardized, programmable, and clinically scalable therapeutic platforms. Future research should focus on improving mechanistic understanding, manufacturing consistency, and clinical validation to facilitate routine clinical application.
Artificial intelligence (AI) and machine learning will significantly accelerate exosome development by optimizing donor-cell selection, cargo loading, biodistribution prediction, manufacturing quality control, and patient-specific treatment strategies. Integration of multi-omics technologies, including genomics, transcriptomics, proteomics, lipidomics, and metabolomics, will further improve biomarker discovery, therapeutic target identification, and personalized exosome design.
Genome-editing technologies such as CRISPR/Cas are expected to expand the therapeutic applications of engineered exosomes by enabling precise correction of disease-associated genes while reducing the limitations associated with viral vectors. In addition, development of hybrid and biomimetic exosomes combining natural extracellular vesicles with synthetic nanomaterials may improve targeting efficiency, cargo capacity, stability, and controlled drug release.
Future exosomes may also become stimulus-responsive "smart" nanocarriers capable of releasing therapeutic cargo selectively in response to pathological conditions such as inflammation, oxidative stress, hypoxia, or pH changes, thereby increasing therapeutic precision and minimizing off-target effects. Integration with injectable hydrogels, biodegradable scaffolds, and advanced biomaterials may further enhance sustained local delivery and tissue regeneration in neurological disorders.
Large-scale commercialization will require automated GMP manufacturing, standardized purification methods, real-time quality monitoring, validated potency assays, and internationally harmonized regulatory guidelines. At the same time, well-designed multicenter clinical trials with long-term follow-up are essential to establish safety, efficacy, and cost-effectiveness in diverse patient populations.
Overall, continued integration of AI, genome editing, biomaterials, multi-omics, and precision medicine is expected to transform engineered exosomes from experimental drug carriers into intelligent therapeutic platforms for the diagnosis, monitoring, and treatment of neurological disorders. Overcoming current scientific, manufacturing, and regulatory challenges will be critical for their successful clinical translation and widespread adoption.
11. CONCLUSION
Engineered exosomes have emerged as promising nanocarriers for the diagnosis and treatment of neurological disorders owing to their excellent biocompatibility, low immunogenicity, intrinsic ability to cross the blood–brain barrier (BBB), and capacity to transport diverse therapeutic cargo. Their natural role in intercellular communication, combined with advances in surface engineering and cargo loading, has enabled the development of targeted delivery systems for proteins, nucleic acids, small-molecule drugs, and gene-editing tools. In addition, exosome-derived biomarkers show considerable potential for the early diagnosis, disease monitoring, and prognosis of major neurological disorders.
Preclinical studies have demonstrated encouraging therapeutic effects in Alzheimer's disease, Parkinson's disease, glioblastoma, ischemic stroke, multiple sclerosis, amyotrophic lateral sclerosis, Huntington's disease, traumatic brain injury, and spinal cord injury. Engineered exosomes have been shown to reduce neuroinflammation, promote neuronal survival, enhance angiogenesis and neurogenesis, improve synaptic plasticity, and facilitate functional recovery. Surface modification with brain-targeting ligands and advances in donor-cell engineering have further improved delivery efficiency and therapeutic specificity, highlighting their potential as next-generation precision nanomedicine.
Despite these advances, several challenges continue to limit clinical translation. Biological heterogeneity, lack of standardized isolation and characterization methods, variable cargo-loading efficiency, incomplete understanding of biodistribution and pharmacokinetics, manufacturing variability, and limited long-term safety data remain major obstacles. Furthermore, scalable GMP-compliant production, robust quality-control systems, validated potency assays, and harmonized international regulatory guidelines are still under development. Addressing these issues is essential for ensuring reproducibility, safety, and regulatory approval.
Future progress will rely on integrating engineered exosomes with artificial intelligence, multi-omics technologies, genome editing, biomaterials, and precision medicine. These interdisciplinary approaches are expected to improve donor-cell selection, optimize therapeutic cargo, enhance targeting accuracy, support large-scale manufacturing, and enable personalized treatment strategies. Equally important are well-designed multicenter clinical trials with standardized protocols and long-term follow-up to establish clinical efficacy and safety.
In conclusion, engineered exosomes represent a transformative platform for brain-targeted drug delivery and neurological therapeutics. Although significant scientific, manufacturing, and regulatory challenges remain, continued advances in exosome engineering, translational research, and clinical validation are expected to accelerate their transition from experimental systems to clinically approved therapies. With ongoing multidisciplinary collaboration, engineered exosomes have the potential to become an integral component of future precision neurology and regenerative medicine.
REFERENCES:


Yash Kothikar*, Tanveer Patel, Engineered Exosomes for Neurological Disorders: From Biomarkers to Brain-Targeted Drug Delivery and Clinical Translation, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 7, 1513-1552. https://doi.org/10.5281/zenodo.21245514
 10.5281/zenodo.21245514
10.5281/zenodo.21245514