
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
1MBBS Third Year, Fergana Medical Institute of Public health Uzbekistan.
2BSC Nursing Fourth year,Dr.Rajendra Gode Nursing Institute of Buldhana, Maharashtra, India.
The emergence of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) catalysed the most rapid vaccine development effort in modern history, culminating in the deployment of messenger RNA (mRNA), viral-vector, recombinant-protein, and inactivated whole-virus vaccines within twelve months of pathogen identification [1–4]. These first-generation vaccines substantially reduced severe disease and mortality, yet their effectiveness has been progressively eroded by waning humoral immunity, the continuous emergence of antigenically divergent variants of concern (VOCs), and persistent inequities in global access [5–9]. This review provides an integrated overview of the immunological, technological, and translational landscape of next-generation COVID-19 vaccines. We critically appraise the design principles of mRNA platforms and their lipid-nanoparticle (LNP) delivery systems, examine emerging modalities including self-amplifying RNA (saRNA), DNA vaccines, virus-like particles (VLPs), and protein-nanoparticle scaffolds, and analyse strategies aimed at achieving pan-sarbecovirus or universal coronavirus immunity through conserved-epitope targeting, mosaic nanoparticle display, and structure-guided immunogen engineering [10–18]. The contributions of nanotechnology to antigen stability, mucosal delivery, and targeted immune activation, together with the role of artificial intelligence (AI) and computational biology in epitope prediction and immunogen optimisation, are discussed in detail [19–25]. We further synthesise comparative real-world effectiveness data for variant-adapted boosters, vaccination considerations in pregnancy, immunocompromised, and paediatric populations, and the health-economic evidence underpinning booster policy, before addressing vaccine safety, regulatory science, global equity, and pandemic preparedness—including the Coalition for Epidemic Preparedness Innovations' 100 Days Mission—and outlining a roadmap toward variant-proof, broadly protective, and globally deliverable coronavirus vaccines [26–30,101–127]. Achieving durable, broadly cross-reactive, and mucosally active immunity will require the convergence of structural vaccinology, advanced delivery science, AI-driven design, and equitable manufacturing—pillars upon which next-generation coronavirus vaccinology must be built.
Coronavirus disease 2019 (COVID-19), caused by SARS-CoV-2, has resulted in more than 770 million confirmed infections and nearly 7 million reported deaths worldwide, while indirect mortality estimates exceed 20 million [1,2]. The unprecedented socioeconomic impact of the pandemic underscored the strategic importance of vaccines as the principal countermeasure against emerging respiratory pathogens [3]. Within twelve months of the publication of the SARS-CoV-2 genome, multiple vaccine platforms received emergency use authorisation, with mRNA vaccines BNT162b2 and mRNA-1273 demonstrating efficacies exceeding 94% against symptomatic disease in pivotal Phase III trials [4,5].
Despite this success, the subsequent emergence of antigenically distinct VOCs—Alpha, Beta, Gamma, Delta, and most notably the Omicron lineage and its descendants (BA.2, BA.5, XBB, JN.1, KP.3)—has progressively eroded neutralising antibody responses and reduced vaccine effectiveness against infection and transmission [6–10]. Waning humoral immunity within 4–6 months of vaccination, immune imprinting effects, and limited mucosal protection at the respiratory portal of entry further constrain current vaccines [11–14]. These limitations, combined with cold-chain dependencies and persistent inequities in vaccine access across low- and middle-income countries (LMICs), have motivated intensive development of next-generation vaccines aimed at broader, more durable, and more equitably deliverable protection [15–18].
This review synthesises the current state of COVID-19 vaccinology and critically examines the scientific, technological, and translational pathways toward next-generation and ultimately universal coronavirus vaccines. We focus on mRNA and saRNA platforms, nanoparticle and VLP scaffolds, conserved-epitope and mosaic-immunogen strategies, mucosal delivery systems, and the integration of AI-driven immunogen design [19–25]. Safety, equity, and pandemic-preparedness dimensions are also addressed [26–30].
2. Overview of Current COVID-19 Vaccines
Four principal vaccine platforms have been deployed at scale during the COVID-19 pandemic: lipid-nanoparticle (LNP)–formulated mRNA vaccines, replication-defective adenoviral-vector vaccines, recombinant spike protein subunit vaccines, and inactivated whole-virion vaccines [4,5,31–34]. Each platform presents distinct immunological profiles, manufacturing constraints, and logistical requirements.
2.1. mRNA Vaccines
The mRNA vaccines BNT162b2 (Pfizer-BioNTech) and mRNA-1273 (Moderna) encode the prefusion-stabilised SARS-CoV-2 spike (S-2P) glycoprotein and are encapsulated within ionisable LNPs that mediate cytosolic delivery and intrinsic adjuvanticity [4,5,31]. Phase III trials reported efficacies of 95% and 94.1%, respectively, against symptomatic COVID-19 in adults [4,5]. Real-world effectiveness studies have confirmed substantial reductions in hospitalisation and death, even against Omicron sublineages, although protection against infection declines markedly within months [6,10,11].
2.2. Viral-Vector Vaccines
Replication-defective adenoviral vectors—including ChAdOx1 nCoV-19 (AZD1222), Ad26.COV2.S, and Sputnik V (rAd26/rAd5)—deliver DNA encoding the spike antigen to host cells [32,33,35]. These vaccines induce robust humoral and cellular responses with simpler cold-chain requirements than mRNA vaccines, but anti-vector immunity and rare thrombosis-with-thrombocytopenia syndrome (TTS) events have limited their continued use in several jurisdictions [36,37].
2.3. Protein Subunit Vaccines
Recombinant subunit vaccines such as NVX-CoV2373 (Novavax) present trimeric prefusion-stabilised spike on nanoparticle scaffolds, formulated with the saponin-based Matrix-M™ adjuvant [34,38]. They combine high immunogenicity with established manufacturing infrastructure and conventional cold-chain compatibility, and have demonstrated efficacies of approximately 90% against ancestral strains [34]. Adapted variant-matched subunit boosters have shown strong cross-neutralisation of contemporary Omicron descendants [38,39].
2.4. Inactivated Vaccines
Inactivated whole-virion vaccines including CoronaVac (Sinovac) and BBIBP-CorV (Sinopharm) have been widely deployed in LMICs [40,41]. They present the complete viral antigen repertoire and are stable at refrigerator temperatures, but elicit comparatively lower neutralising antibody titres and require multiple boosters to maintain protection against VOCs [41,42].
3. Limitations of First-Generation Vaccines
Although first-generation COVID-19 vaccines have averted millions of deaths, four major limitations have driven the search for next-generation platforms: (i) waning humoral immunity, (ii) variant-driven immune escape, (iii) limited mucosal protection, and (iv) cold-chain and equity constraints [6–18,42].
Serological studies demonstrate a marked decline in neutralising antibody titres within 4–6 months of primary vaccination, with concomitant reductions in vaccine effectiveness against symptomatic infection [11,12]. Antigenic drift across the receptor-binding domain (RBD) of the spike protein—particularly in Omicron and its descendants XBB.1.5, EG.5, BA.2.86, JN.1, and KP.3—has dramatically reduced neutralising titres elicited by ancestral-strain vaccines [7–10,43]. Furthermore, intramuscular vaccines induce primarily systemic IgG responses with minimal mucosal IgA, leaving the upper respiratory tract relatively unprotected and permitting onward transmission [13,14,44]. Finally, ultra-cold storage requirements for first-generation mRNA vaccines (–70 °C for BNT162b2; –20 °C for mRNA-1273), short shelf-lives once thawed, and high per-dose costs have constrained equitable deployment in LMICs [15–18,45].
4. mRNA Vaccine Platforms
mRNA vaccines comprise an in vitro–transcribed, 5′-capped, polyadenylated, nucleoside-modified mRNA encoding a target antigen, encapsulated within ionisable LNPs composed of an ionisable cationic lipid, a phospholipid, cholesterol, and a polyethylene glycol (PEG)–lipid [31,46,47]. Following intramuscular injection, LNPs are internalised by antigen-presenting cells, the mRNA escapes from endosomes into the cytosol, and the encoded antigen is translated, processed, and presented on both MHC class I and II molecules, eliciting balanced humoral and cellular immunity [46–48].
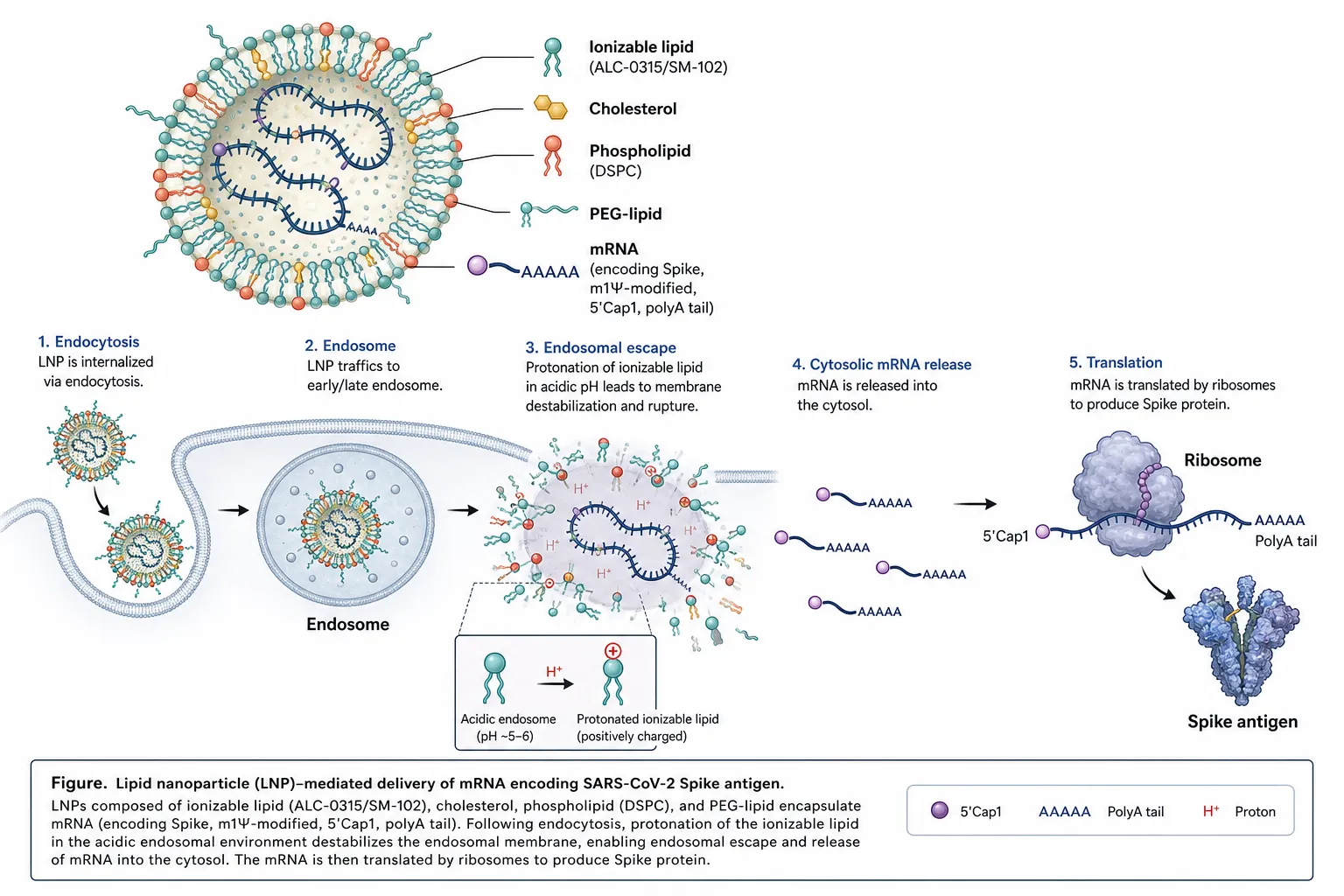
Figure 1. Lipid nanoparticle (LNP)–mediated delivery of mRNA encoding the SARS-CoV-2 Spike antigen. Ionisable lipids (ALC-0315/SM-102), cholesterol, phospholipid (DSPC) and PEG-lipid encapsulate m1Ψ-modified, 5′-Cap1, polyadenylated mRNA. Following endocytosis, protonation of the ionisable lipid in the acidic endosome drives endosomal escape; cytosolic mRNA is translated by ribosomes into Spike antigen.
4.1. Nucleoside Modification and Translation Efficiency
Incorporation of N1-methylpseudouridine (m1Ψ) markedly reduces innate immune sensing by Toll-like receptors 7/8 and RIG-I, decreases interferon-driven translational shutdown, and enhances protein expression compared with unmodified mRNA [46,49]. Codon optimisation, optimised 5′ and 3′ untranslated regions, and Cap1 structures further improve translational efficiency [47,50].
4.2. Lipid Nanoparticles as Delivery Vehicles
Ionisable lipids such as ALC-0315 and SM-102 are protonated within the acidic endosomal environment, facilitating membrane disruption and cytosolic mRNA release [47,51]. PEG-lipids stabilise the LNP and modulate biodistribution, although anti-PEG antibodies have been implicated in rare hypersensitivity reactions [52]. Next-generation LNPs incorporate biodegradable ester linkages, reduced PEG content, and tissue-targeting ligands to improve safety, stability, and organ specificity [51,53].
4.3. Advantages and Limitations
mRNA platforms offer rapid design-to-clinic timelines (weeks rather than months), platform reusability across pathogens, scalable cell-free manufacturing, and absence of integration risk [46–48]. Limitations include cold-chain dependency, batch-to-batch heterogeneity, LNP-related reactogenicity, and the requirement for repeated boosting [52,53].
5. Emerging Vaccine Technologies
Beyond conventional mRNA platforms, several emerging modalities are advancing through preclinical and clinical pipelines.
5.1. Self-Amplifying RNA (saRNA) Vaccines
saRNA vaccines, derived from alphavirus replicons, encode both the antigen of interest and a viral RNA-dependent RNA polymerase, enabling intracellular amplification of antigen-encoding transcripts and substantially reducing the required dose [54,55]. ARCT-154 (Arcturus/CSL) became the first saRNA COVID-19 vaccine licensed for human use in Japan in 2023 [56].
5.2. DNA Vaccines
Plasmid DNA vaccines such as ZyCoV-D (Cadila) are thermostable, inexpensive to manufacture, and administered via needle-free intradermal devices [57]. Although immunogenicity is generally lower than mRNA vaccines, electroporation and improved plasmid designs are narrowing this gap [57,58].
5.3. Virus-Like Particles and Protein Nanoparticles
Virus-like particles (VLPs) and self-assembling protein nanoparticles (e.g., ferritin, I53-50, mi3) display multivalent spike or RBD antigens in a highly ordered repetitive array, enhancing B-cell receptor cross-linking and germinal-centre responses [16,17,59]. Computationally designed nanoparticles such as SKYCovione (GBP510) have demonstrated superior immunogenicity in head-to-head clinical trials [60].
5.4. Mucosal and Intranasal Vaccines
Intranasal and inhaled vaccines based on adenoviral vectors, live-attenuated viruses, or protein-nanoparticle formulations aim to induce sterilising mucosal immunity through secretory IgA and tissue-resident memory T cells in the respiratory tract [14,44,61,62]. Several candidates (e.g., iNCOVACC, Convidecia Air) have received regulatory approval, although correlates of mucosal protection remain to be standardised [61,62].
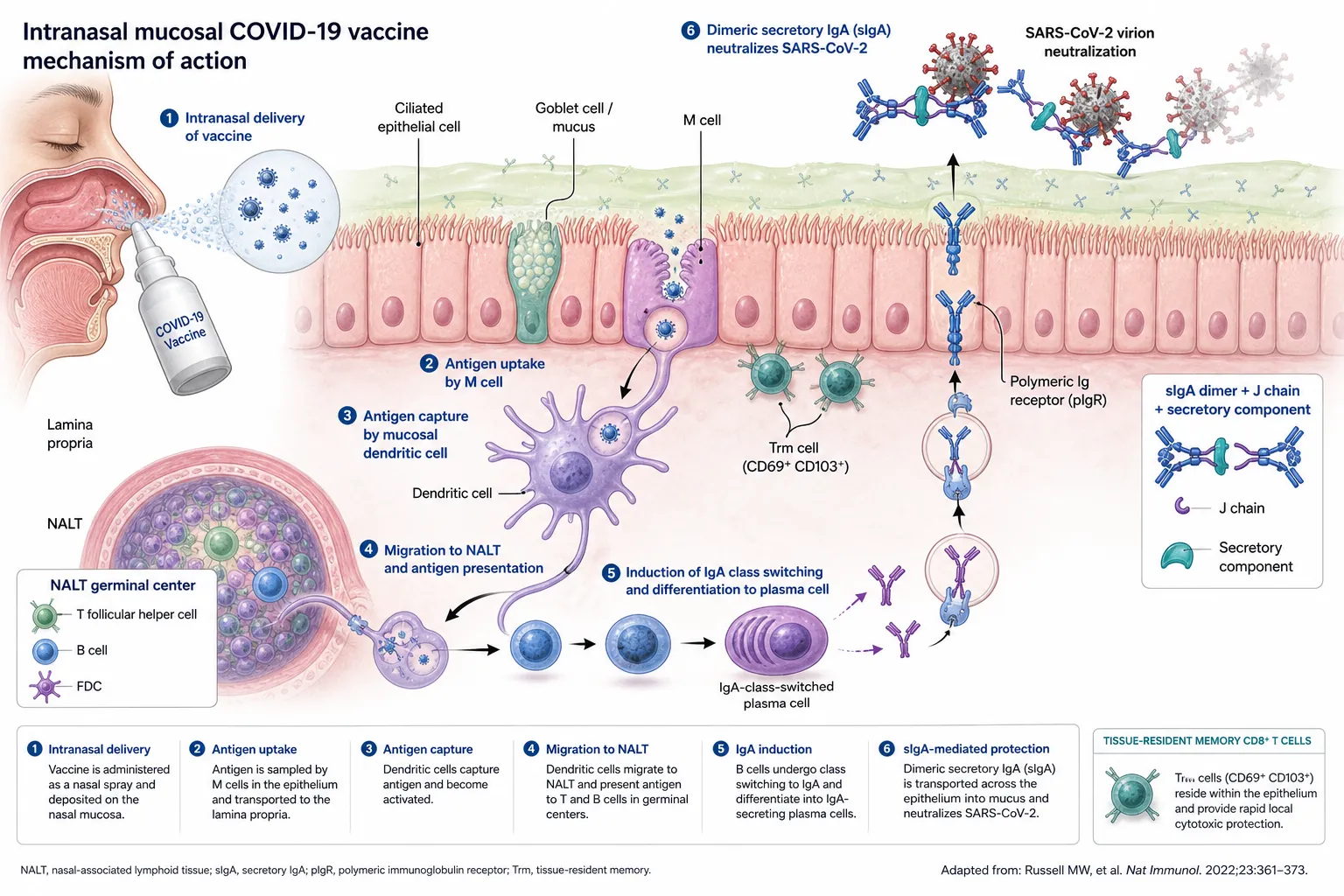
Figure 5. Mechanism of action of an intranasal mucosal COVID-19 vaccine. Antigen sampled by M cells is captured by mucosal dendritic cells and presented in NALT germinal centres, driving IgA class-switching and differentiation into plasma cells. Dimeric IgA, complexed with the J chain and secretory component, is transported across the epithelium via pIgR and neutralises SARS-CoV-2 in the mucus layer. Tissue-resident memory CD8⁺ T cells (CD69⁺CD103⁺) provide rapid local cytotoxic protection.
6. Immunological Basis of Vaccine Protection
Effective vaccine-induced immunity requires coordinated activation of innate and adaptive arms of the immune system [63]. LNP and adjuvant components engage pattern-recognition receptors on dendritic cells, driving their maturation, migration to draining lymph nodes, and antigen presentation to naïve T cells [46,63]. CD4⁺ T follicular helper (Tfh) cells support germinal-centre B-cell affinity maturation and class switching, producing high-affinity neutralising IgG antibodies, while CD8⁺ cytotoxic T lymphocytes eliminate infected cells [64,65].
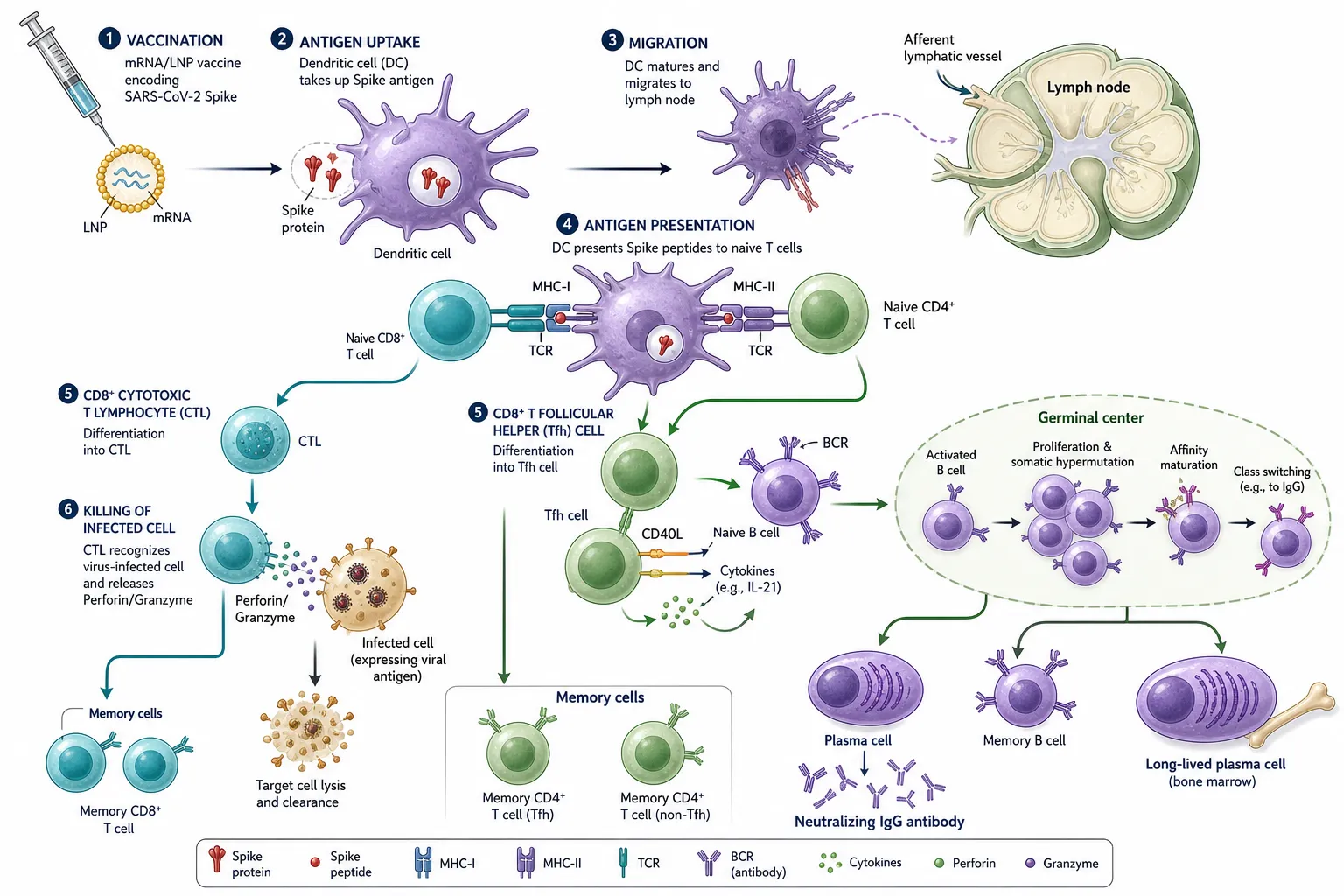
Figure 2. Vaccine-induced adaptive immune response. Dendritic cells acquire Spike antigen, migrate to the draining lymph node, and present peptides on MHC-I and MHC-II to naïve CD8⁺ and CD4⁺ T cells. CD4⁺ Tfh cells license germinal-centre B cells to undergo affinity maturation and class-switching, generating neutralising IgG-secreting plasma cells, long-lived plasma cells, and memory B/T cells; CD8⁺ CTLs eliminate infected cells via perforin/granzyme.
Long-lived plasma cells in the bone marrow sustain serum antibody titres, and memory B and T cells underpin anamnestic responses upon reinfection [65,66]. Importantly, T-cell epitopes are substantially more conserved across SARS-CoV-2 variants than B-cell neutralising epitopes, contributing to preserved protection against severe disease despite antibody evasion by Omicron-lineage variants [66,67]. Mucosal protection is mediated by secretory dimeric IgA and tissue-resident memory CD8⁺ T cells (Trm), which are inefficiently induced by intramuscular vaccination [14,44,61].
7. SARS-CoV-2 Variants and Immune Escape
SARS-CoV-2 evolution has been characterised by punctuated emergence of antigenically distinct lineages bearing convergent mutations in the spike RBD and N-terminal domain that confer immune escape, altered receptor binding, and modified transmissibility [6–10,43,68]. Key escape mutations include K417N/T, L452R, E484K/A, N501Y, and the extensively mutated Omicron RBD, with continued diversification in BA.2.86, JN.1, and KP.3 sublineages [8–10,43].
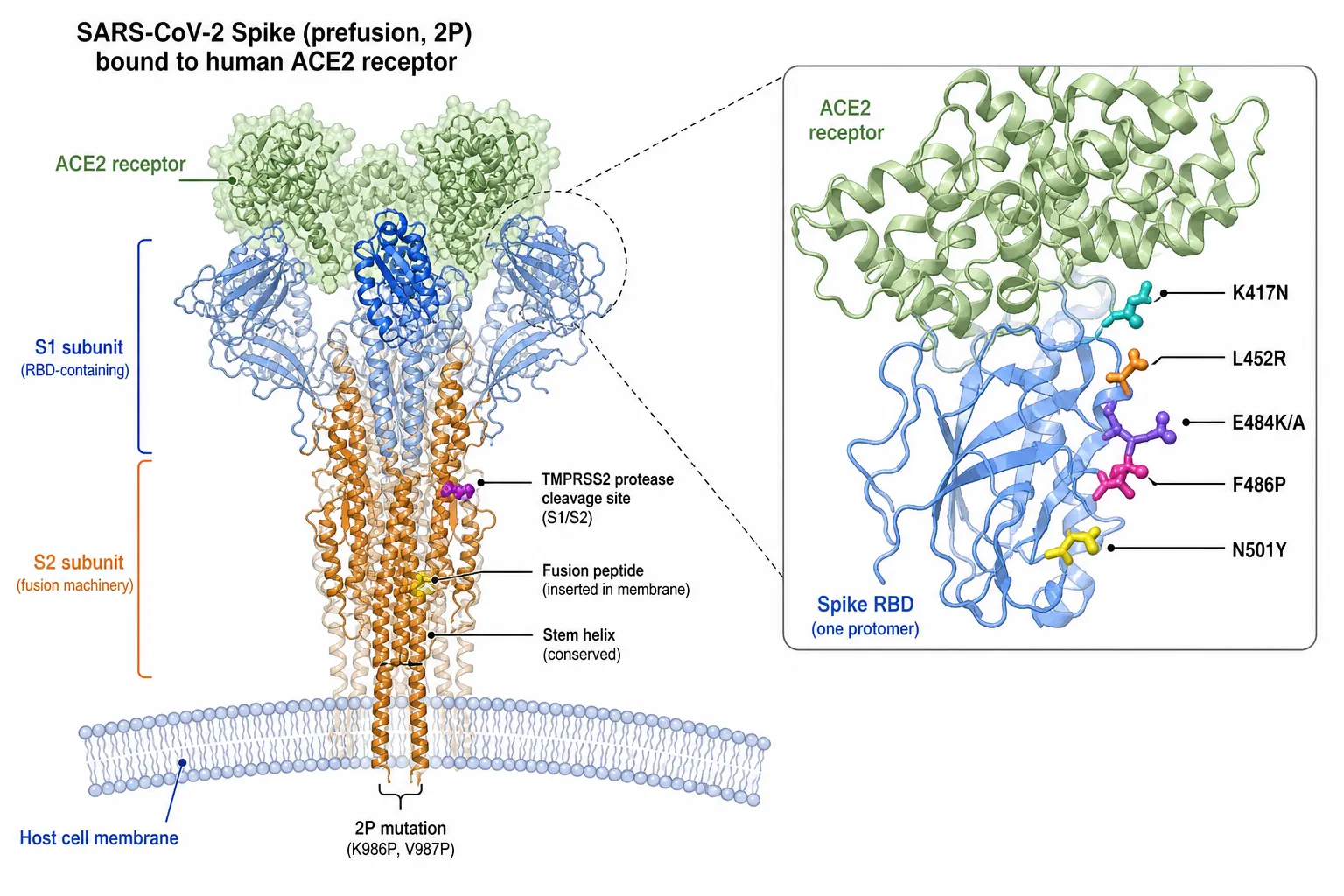
Figure 3. Prefusion-stabilised (2P) SARS-CoV-2 Spike trimer engaging human ACE2. Inset highlights key receptor-binding domain (RBD) immune-escape residues (K417N, L452R, E484K/A, F486P, N501Y), the S1/S2 TMPRSS2 cleavage site, the fusion peptide, and the conserved stem-helix region of the S2 subunit.
Antigenic cartography demonstrates that contemporary Omicron descendants occupy antigenic space far removed from the ancestral Wuhan-Hu-1 strain, necessitating periodic reformulation of vaccines analogous to seasonal influenza updates [43,68]. Bivalent and monovalent Omicron-adapted boosters (BA.4/5, XBB.1.5, JN.1) have restored neutralising activity against circulating strains, but durability and breadth remain suboptimal [38,39,69].
8. Universal Coronavirus Vaccine Strategies
A universal coronavirus vaccine—capable of eliciting broadly protective immunity across SARS-CoV-2 variants, sarbecoviruses, and potentially all betacoronaviruses—has emerged as a strategic priority for pandemic preparedness [18,70–73]. Three principal strategies are under investigation.
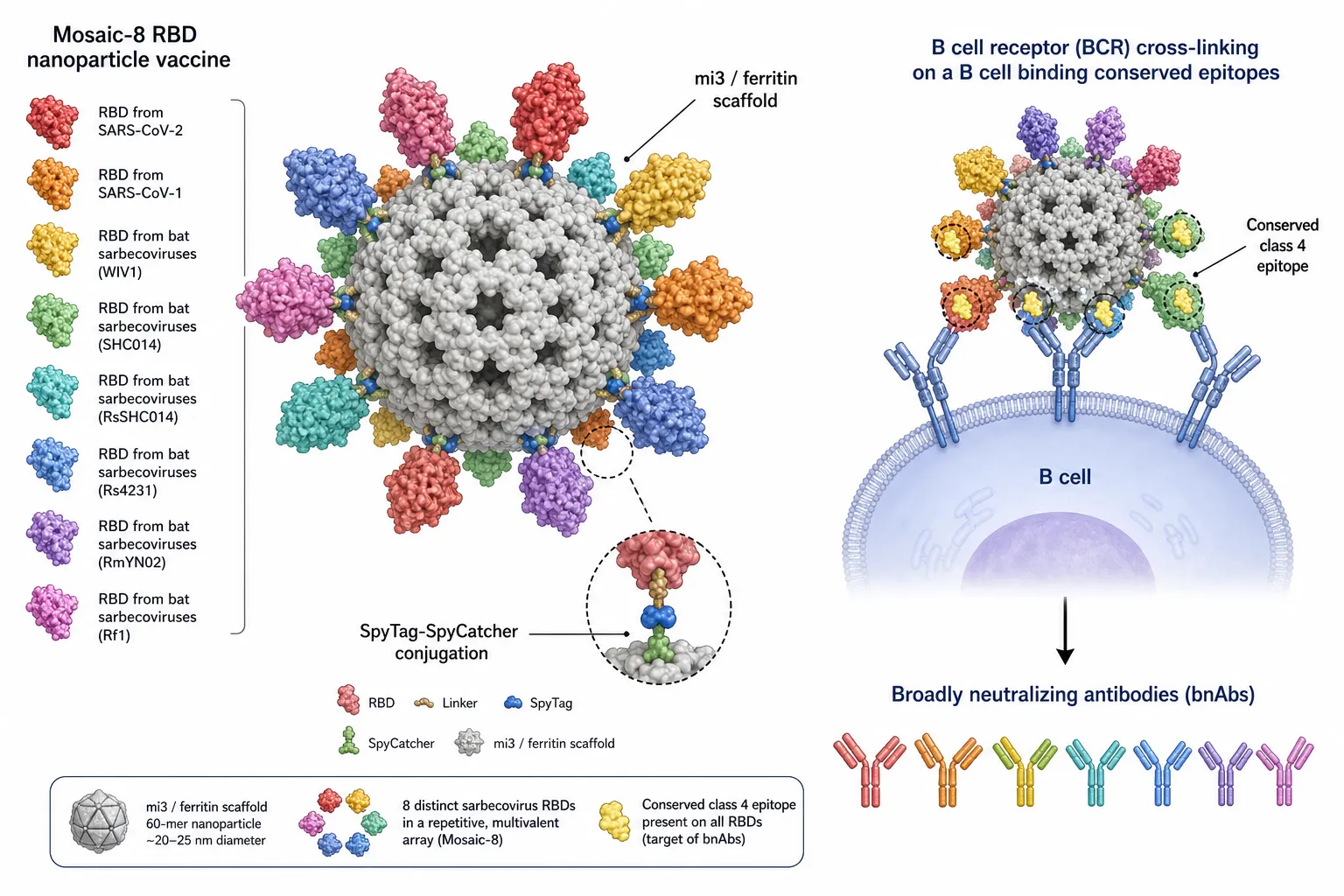
Figure 4. Mosaic-8 RBD nanoparticle vaccine. Eight distinct sarbecovirus RBDs (SARS-CoV-2, SARS-CoV-1, WIV1, SHC014, RsSHC014, Rs4231, RmYN02, Rf1) are conjugated to an mi3/ferritin 60-mer scaffold via SpyTag–SpyCatcher chemistry. Multivalent display focuses B-cell receptor cross-linking on the conserved class-4 epitope shared across RBDs, eliciting broadly neutralising antibodies (bnAbs).
8.1. Conserved-Epitope Targeting
Structure-guided immunogens focus B- and T-cell responses on conserved regions including the spike stem-helix, fusion peptide, and S2 subunit, as well as conserved RBD class 4 epitopes that elicit broadly neutralising antibodies (bnAbs) [70,71,74].
8.2. Mosaic Nanoparticle Vaccines
Mosaic-8 and mosaic-RBD nanoparticles co-display RBDs from multiple sarbecoviruses on a single mi3 or ferritin scaffold, focusing the antibody response on conserved epitopes and eliciting cross-protective immunity in non-human primates [16,72,73].
8.3. Chimeric and Computationally Designed Immunogens
Chimeric spike immunogens, resurfaced RBDs, and germline-targeting probes designed via computational protein engineering aim to recapitulate the maturation pathways of bnAbs identified from convalescent donors [22,75,76].
9. Role of Nanotechnology in Next-Generation Vaccines
Nanotechnology underpins virtually every next-generation vaccine modality, providing platforms for antigen multivalency, controlled release, targeted delivery, and immune modulation [19,77]. LNPs, polymeric nanoparticles (PLGA, chitosan), inorganic nanoparticles (gold, silica), and self-assembling protein nanoparticles each offer distinct physicochemical and immunological profiles [19,77,78]. Surface functionalisation with antigen-presenting cell–targeting ligands (e.g., mannose, anti-DEC-205, anti-CLEC9A) enhances dendritic-cell uptake and cross-presentation [79]. Thermostable nanoparticle formulations, lyophilised mRNA-LNPs, and microneedle patches are being developed to overcome cold-chain limitations and enable equitable global distribution [80,81].
10. Artificial Intelligence and Computational Vaccine Design
Artificial intelligence (AI) and machine learning have become indispensable to modern vaccinology, accelerating antigen discovery, epitope prediction, and immunogen optimisation [20–25,82]. Deep-learning models such as NetMHCpan-4.1 and MHCflurry predict peptide–MHC binding with high accuracy, enabling rational selection of broadly conserved T-cell epitopes [21,83]. AlphaFold2 and RoseTTAFold have transformed structural vaccinology by enabling reliable prediction of antigen conformations, facilitating prefusion stabilisation, glycan engineering, and de novo immunogen design [22,84].
Generative AI models including ProGen, RFdiffusion, and ESMFold are being applied to design novel protein scaffolds for nanoparticle vaccines and to engineer immunogens that focus antibody responses on conserved epitopes [22,75,85]. AI-driven pipelines integrating genomic surveillance, structural prediction, and immunoinformatics now enable near-real-time vaccine reformulation in response to emerging variants [25,86].
11. Safety and Adverse Effects
First-generation COVID-19 vaccines have an established safety profile supported by post-authorisation surveillance of billions of administered doses [26,87,88]. Most adverse events are mild and transient, including injection-site pain, fatigue, headache, and low-grade fever [87]. Rare but clinically significant events include myocarditis and pericarditis, predominantly in young males following mRNA vaccination (incidence ~1–10 per 100,000 doses) [89,90]; thrombosis with thrombocytopenia syndrome (TTS) following adenoviral-vector vaccines [36,37]; and anaphylaxis associated with PEG components of LNPs [52,91]. Long-term surveillance studies have not identified causal links between vaccination and post-acute sequelae [26,88]. Next-generation vaccines aim to retain immunogenicity while reducing reactogenicity through optimised LNP composition, lower antigen doses (enabled by saRNA), and mucosal delivery [54,61,92].
12. Global Vaccine Equity and Distribution
Global COVID-19 vaccine deployment has been marked by profound inequities: by mid-2022, more than 70% of high-income populations had received at least one dose, compared with fewer than 20% in many low-income countries [15,93,94]. Bilateral procurement, intellectual-property restrictions, manufacturing concentration, and cold-chain limitations have all contributed to this disparity [15,93]. Initiatives such as COVAX, the mRNA technology-transfer hub in South Africa, and regional manufacturing partnerships represent essential structural responses [94,95]. Thermostable formulations, microneedle patches, single-dose regimens, and intranasal vaccines are technological levers that can further reduce delivery costs and improve coverage in resource-limited settings [80,81,96].
13. Comparative Clinical Effectiveness of Variant-Adapted Boosters
The repeated reformulation of mRNA vaccines to match circulating Omicron sublineages has generated a substantial real-world evidence base on comparative booster effectiveness. Monovalent XBB.1.5-adapted vaccines, introduced for the 2023–2024 immunisation season, were evaluated across multiple national surveillance networks before any randomised efficacy trial had reported, making observational target-trial and test-negative designs the principal source of effectiveness data [101,102].
A systematic review and meta-analysis pooling more than fifty million participants across twenty-one studies found that XBB.1.5-adapted vaccines conferred meaningfully greater protection against severe outcomes than against infection itself, a pattern consistent with earlier-generation boosters and attributable to the relative durability of cellular over humoral immunity [103]. Effectiveness against hospitalisation was higher during periods of antigenic match with circulating XBB-lineage viruses than after the emergence of the more antigenically distant BA.2.86/JN.1 lineage, reaffirming that the magnitude of antigenic distance between vaccine strain and circulating strain remains a principal determinant of short-term protection [104,105]. A United States Veterans Health Administration target-trial emulation similarly documented effectiveness against infection that was highest in the first ten weeks after vaccination and declined progressively thereafter, with protection against hospitalisation proving more durable than protection against infection across the six-month follow-up period [102]. A multinational European surveillance network reported that, among adults aged sixty-five years and older, hospitalisation rates converged between vaccinated and unvaccinated cohorts within approximately eight to nine months of the autumn 2023 campaign, coinciding with the JN.1 wave and underscoring the practical case for at least annual reformulation in older adults [106].
Table 1. Reported effectiveness of monovalent XBB.1.5-adapted mRNA boosters against SARS-CoV-2 outcomes, 2023–2024 season.
|
Outcome |
Effectiveness (95% CI) |
Evaluation window |
Source |
|
Symptomatic SARS-CoV-2 infection |
52.9% (47.6–57.6%) |
First month post-dose |
Pooled meta-analysis, 21 studies [103] |
|
COVID-19-related hospitalisation |
64.4% (59.3–68.9%) |
First month post-dose |
Pooled meta-analysis, 21 studies [103] |
|
COVID-19-related death |
77.3% (67.1–84.3%) |
First month post-dose |
Pooled meta-analysis, 21 studies [103] |
|
Hospitalisation, XBB-lineage |
54.2% (36.1–67.1%) |
7–89 days post-dose |
IVY Network, 26 US hospitals [104] |
|
Hospitalisation, JN-lineage |
32.7% (1.9–53.8%) |
7–89 days post-dose |
IVY Network, 26 US hospitals [104] |
|
Hospitalisation or death, ≥65 years |
Converged with unvaccinated by ~8–9 months |
Autumn 2023–Summer 2024 |
VEBIS EU/EEA network [106] |
CI, confidence interval. Effectiveness estimates are drawn from heterogeneous study designs (test-negative case-control, target-trial emulation, and pooled meta-analysis) and are not directly comparable across rows; they are presented together to illustrate the consistent gradient of higher protection against severe disease than against infection, and the attenuation of protection against the antigenically more distant JN.1 lineage.
Taken together, these data support three inferences relevant to next-generation vaccine design. First, the gradient of protection—lowest against infection, intermediate against hospitalisation, and highest against death—implies that current platforms are better suited to preventing severe outcomes than to achieving sterilising immunity, reinforcing the rationale for mucosal and saRNA approaches discussed in Sections 4 and 5 [103,104]. Second, the sensitivity of effectiveness to antigenic distance between vaccine and circulating strains argues for faster, more frequent reformulation cycles or, alternatively, for the conserved-epitope and mosaic-nanoparticle strategies discussed in Section 8, which are explicitly designed to be insensitive to this antigenic drift [105,107]. Third, the consistent finding that protection in older adults wanes within six to nine months provides the empirical basis for the six-monthly booster recommendations now in place in several high-income countries [101,106].
14. Vaccination in Special Populations
14.1. Pregnancy
Pregnant individuals face an elevated risk of severe COVID-19 and adverse perinatal outcomes following SARS-CoV-2 infection, yet vaccine uptake in this group has lagged behind the general population in most countries, reflecting both historical exclusion from pre-licensure trials and persistent safety concerns among patients and clinicians [108,109]. A meta-analysis of twenty-three studies encompassing more than 117,000 vaccinated pregnant individuals found that mRNA vaccination was approximately 90% effective against confirmed infection from the seventh day after the second dose, and was associated with a significantly lower risk of stillbirth relative to the unvaccinated cohort [108]. No excess risk of miscarriage, preterm birth, placental abruption, postpartum haemorrhage, or neonatal intensive care admission was identified across the pooled studies, and subsequent real-world cohorts evaluating effectiveness against infection and hospitalisation have reported concordant findings [108,110]. Maternal vaccination further confers passive protection to the neonate through transplacental and breast-milk transfer of anti-spike immunoglobulin, an effect that is most pronounced when vaccination occurs in the second or third trimester [108]. These findings have informed professional society guidance recommending COVID-19 vaccination at any stage of pregnancy, although ongoing prospective pharmacovigilance remains necessary to characterise effectiveness against newer variants and to refine guidance on optimal trimester-specific timing [109,110].
14.2. Immunocompromised Individuals
Solid-organ transplant recipients, haematopoietic stem-cell transplant recipients, and patients receiving B-cell-depleting or other immunosuppressive therapies mount substantially attenuated humoral responses to standard COVID-19 vaccine schedules. A meta-analysis of twenty-six studies comprising more than three thousand immunocompromised patients found that the likelihood of seroconversion after a two-dose mRNA primary series was approximately 48% lower than in immunocompetent controls, with transplant recipients showing the most pronounced impairment and patients with autoimmune disease or solid-organ malignancy showing comparatively preserved responses [111]. An additional vaccine dose administered as an extended primary series produces a meaningful, if incomplete, improvement in seroconversion: a rapid review of randomised and observational studies found that median antibody response rates rose from 41% after the standard primary series to 67% after a supplementary dose, without a corresponding increase in serious adverse events [112]. Comparative data on heterologous versus homologous additional dosing in non-seroconverted immunocompromised patients have not demonstrated a clear immunogenicity advantage for either approach, although heterologous viral-vector dosing was associated with a modestly higher incidence of headache [113,114]. These findings underpin the current World Health Organization and national guidance recommending a three-dose extended primary series, rather than a conventional two-dose series followed by a booster, for moderately to severely immunocompromised individuals [113].
14.3. Paediatric Populations
Paediatric COVID-19 vaccination programmes have prioritised prevention of multisystem inflammatory syndrome in children and severe acute disease over interruption of transmission, reflecting the markedly lower baseline risk of severe outcomes in children relative to older adults [44,108]. Age-de-escalated mRNA dosing regimens (typically one-third to one-tenth of the adult antigen mass) have demonstrated immunogenicity and reactogenicity profiles broadly comparable to those observed in young adults, although real-world effectiveness data in children remain less extensive than in adult cohorts, and optimal booster intervals for this age group continue to be refined as additional surveillance data accrue [44].
15. Health-Economic Evaluation of Booster Strategies
As COVID-19 vaccination programmes have transitioned from emergency mass campaigns to recurring seasonal or risk-stratified booster schedules, health-economic evaluation has become central to policy decisions regarding which populations to prioritise and how frequently to recommend revaccination [115,116]. Cost-effectiveness analyses across several high-income health systems converge on a consistent finding: booster vaccination is most economically favourable when targeted at older adults and individuals with significant comorbidities, who carry the greatest absolute risk of hospitalisation and death, while remaining of more marginal economic value in younger, healthier cohorts at prevailing vaccine prices [115,117,118].
Table 2. Selected cost-effectiveness evaluations of COVID-19 booster vaccination programmes.
|
Setting / population |
Strategy evaluated |
ICER (US$/QALY, approx.) |
Source |
|
South Korea, all adults |
Uniform annual booster allocation |
~25,800 |
Transmission-dynamic model [115] |
|
South Korea, adults ≥65 y |
Age-prioritised annual booster |
~13,800 |
Transmission-dynamic model [115] |
|
Qatar, general population |
mRNA primary series, pre-Omicron phase |
~68,500 |
Retrospective cohort + modelling [116] |
|
USA, adults ≥50 y |
Second booster dose |
Cost-saving (dominant) |
SEIR–Markov model [117] |
|
Australia, adults ≥16 y |
mRNA booster, 3 months post-dose 2 |
Below A$50,000 WTP threshold |
Markov decision model [118] |
ICER, incremental cost-effectiveness ratio; QALY, quality-adjusted life year; WTP, willingness-to-pay. Estimates derive from heterogeneous national settings, time horizons, and modelling assumptions and should not be compared directly across rows; “dominant” denotes an intervention that is simultaneously more effective and less costly than the comparator.
A recurring methodological theme across these evaluations is that cost-effectiveness is more sensitive to vaccine acquisition price than to plausible variation in vaccine effectiveness, implying that procurement pricing—particularly for lower- and middle-income settings—exerts a disproportionate influence on the economic case for sustained booster programmes [115,119]. Analyses that incorporate broader societal costs, including productivity losses and long-COVID-attributable morbidity, generally report more favourable cost-effectiveness ratios than analyses confined to direct healthcare-system costs, suggesting that narrowly framed healthcare-perspective evaluations may understate the full economic value of booster vaccination [119,120]. These findings reinforce the case made elsewhere in this review for thermostable, lower-cost next-generation formulations: even modest reductions in per-dose price or cold-chain expenditure could materially improve the economic case for sustained, broadly targeted booster programmes in resource-constrained settings [80,116].
16. Regulatory Pathways and Post-Authorisation Surveillance
The regulatory framework governing variant-adapted COVID-19 vaccines has itself evolved substantially since 2020. Rather than requiring full pre-licensure efficacy trials for each reformulated vaccine, regulators in the United States, European Union, and other jurisdictions have adopted an immunobridging approach analogous to that used for seasonal influenza vaccines, in which strain changes are authorised on the basis of immunogenicity and safety data from a subset of recipients, supplemented by post-authorisation effectiveness and safety surveillance [101,121]. This approach materially shortens the interval between variant emergence and vaccine availability, but it also means that real-world effectiveness data—of the kind summarised in Section 13—necessarily arrive after, rather than before, a reformulated vaccine reaches the public, placing a correspondingly greater burden on robust post-marketing surveillance infrastructure [101,102].
Pharmacovigilance networks established or substantially expanded during the pandemic, including the Vaccine Safety Datalink in the United States and the VEBIS electronic health-record network across seven European countries, have enabled near-real-time monitoring of rare adverse events and effectiveness signals at a scale not previously achievable for any vaccine programme [89,106]. This infrastructure was instrumental in characterising the rare myocarditis signal associated with mRNA vaccination in young males and the thrombosis-with-thrombocytopenia signal associated with adenoviral-vector vaccines discussed in Section 11, in both cases within months of widespread rollout [36,89,90]. Sustaining and further integrating these surveillance networks—ideally linking genomic variant surveillance directly to clinical and immunogenicity data—represents an important structural priority for the next generation of coronavirus vaccine governance, independent of which specific platform or antigen design ultimately predominates [101,121].
17. Pandemic Preparedness Modelling and the 100 Days Mission
The Coalition for Epidemic Preparedness Innovations (CEPI) 100 Days Mission, endorsed by the Group of Seven and Group of Twenty forums, articulates a target of making a safe and effective vaccine available within 100 days of the identification of a novel pandemic pathogen, compared with the approximately 326 days that elapsed between the publication of the SARS-CoV-2 genome and the first emergency-use authorisation of a COVID-19 vaccine [122,123]. The Mission rests on three structural pillars: a library of prototype vaccines pre-developed against representative pathogens from priority viral families, so that a genuinely novel pathogen can be addressed by rapidly substituting its antigen into an already-characterised platform; harmonised regulatory and clinical-trial-readiness frameworks that can be activated immediately upon outbreak declaration; and distributed, regionally diversified manufacturing capacity intended to avoid the bilateral procurement bottlenecks that characterised early COVID-19 vaccine distribution [122,124].
Mathematical modelling of a counterfactual scenario in which 100-day vaccine availability had been achieved during the actual COVID-19 pandemic estimated substantial reductions in global infections and deaths relative to the observed timeline, providing a quantitative rationale for continued investment in prototype-pathogen libraries and rapid chemistry-manufacturing-and-controls frameworks [125]. Implementation efforts since 2022 have included CEPI-coordinated clinical-trial-readiness networks in East and Central Africa, designed to build durable regional research capacity rather than transient, sponsor-driven trial infrastructure, and a dedicated CEPI rapid-response framework spanning manufacturing, formulation, analytics, supply chain, and facilities readiness [124,126]. The mRNA platform features prominently in 100 Days Mission planning because its sequence-based design-to-clinic workflow, described in Section 4, is uniquely suited to rapid antigen substitution; however, planners have emphasised that platform diversity across mRNA, viral-vector, and protein-subunit modalities remains important for manufacturing resilience and for reaching populations with differing logistical or acceptability constraints [122,126].
Analyses examining the 100 Days Mission specifically through the lens of low- and middle-income countries have cautioned that a 100-day global timeline is compatible with persistent within-pandemic inequity if early-authorised doses are absorbed disproportionately by high-income purchasers, echoing the COVAX-era access disparities discussed in Section 12 [123,127]. Survey-based assessments of pandemic-preparedness experts from both high- and low/middle-income countries similarly identify equitable allocation mechanisms, rather than raw development speed alone, as the more uncertain determinant of whether the Mission's benefits will be realised broadly [127]. The adoption of the WHO Pandemic Agreement in May 2025, which embeds equity and collaborative innovation as core principles, represents a parallel governance development intended to address this concern, although its practical effect on dose allocation during a future pandemic remains to be tested [127].
18. FUTURE PERSPECTIVES
The trajectory of coronavirus vaccinology is converging on three strategic objectives: durability, breadth, and equity [18,97]. Durability will require induction of long-lived plasma cells and memory B-cell pools through optimised antigen kinetics, slow-release depot formulations, and rational adjuvant selection [98,99]. Breadth will be achieved through conserved-epitope immunogens, mosaic nanoparticle display, and AI-designed pan-sarbecovirus scaffolds [16,22,72,73]. Equity will depend on thermostable, needle-free, and locally manufacturable vaccines, supported by enduring technology-transfer infrastructure [80,94,95].
Beyond SARS-CoV-2, the same platforms are accelerating vaccine development for influenza, respiratory syncytial virus, HIV, and pandemic-prone pathogens identified by the WHO R&D Blueprint [100]. Continued investment in genomic surveillance, prototype-pathogen vaccine libraries, and rapid-response manufacturing networks is essential to translate the lessons of COVID-19 into durable pandemic preparedness [29,30,100].
CONCLUSION
First-generation COVID-19 vaccines represent a singular achievement of modern biomedicine, yet their limitations—waning immunity, variant escape, limited mucosal protection, and inequitable access—define the imperative for next-generation platforms. The convergence of mRNA and saRNA technologies, computationally designed nanoparticles, mucosal delivery systems, and AI-driven immunogen engineering offers a credible scientific pathway toward durable, broadly cross-reactive, and globally deliverable coronavirus vaccines. Realising this vision will require sustained investment in basic immunology, translational science, equitable manufacturing, and coordinated global governance—pillars that together will determine the resilience of humanity against future coronavirus threats.
Acknowledgements
The authors thank colleagues at the participating institutions for critical discussion of the manuscript.
Funding
This review received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Conflicts of Interest
The authors declare no competing interests.
Author Contributions
All authors contributed to the conceptualisation, literature review, drafting, and critical revision of the manuscript and approved the final version for submission.
REFERENCES


Nandhini Thangavel, Pratiksha Jadhao, Next-Generation COVID-19 Vaccines: From mRNA Platforms to Universal Coronavirus Immunity, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 6, 7731-7751, https://doi.org/10.5281/zenodo.21067094
 10.5281/zenodo.21067094
10.5281/zenodo.21067094