
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
Department of Pharmacology, Shantiniketan College of Pharmacy, Dhotre (Bk.), Tal.: Parner, Dist.: Ahilyanagar-414 304, Maharashtra, India.
Background: Clitoria ternatea L. (butterfly pea; Fabaceae) is a perennial tropical legume with a multi-century history in Ayurvedic medicine. While individual pharmacological activities have been documented, a comprehensive critical synthesis of evidence specifically addressing the concurrent in vitro modulation of oxidative stress and inhibition of protein denaturation — two interlinked pathways underlying chronic disease and inflammation — has not previously been published. Objectives: To critically consolidate published peer-reviewed evidence on the in vitro antioxidant and protein denaturation-inhibitory properties of C. ternatea extracts; to examine phytochemical determinants and molecular mechanisms; and to identify research gaps requiring future investigationMethods: A structured literature search was performed across PubMed, Scopus, and Web of Science. Studies from 2010 to 2025 employing validated in vitro assays including DPPH, FRAP, ABTS, BSA heat denaturation, protein carbonyl, and thiol measurement assays were critically evaluated. Key Findings: Aqueous and hydroethanolic flower extracts demonstrated DPPH IC50 values as low as 0.47 mg/mL, FRAP of 0.38 mmol FeSO4 eq./mg, and 75-80% intracellular ROS suppression in macrophage models. Protein denaturation inhibition studies revealed IC50 values comparable to diclofenac sodium (methanolic leaf extract IC50 = 120.05 µg/mL), with protection from methylglyoxal-induced carbonylation (22% reduction) and thiol depletion (56% prevention). The pyrogallol B-ring structure of delphinidin-based ternatins is identified as the primary structural basis for radical scavenging, while phenolic acid-mediated hydrogen bonding and NF-kB pathway inhibition by quercetin glycosides underpin anti-denaturation activity. Novelty: This review presents the first systematic integration of antioxidant and protein denaturation inhibitory evidence as an interlinked mechanistic framework, proposes a unified dual-mechanism
Oxidative stress — defined as a disruption in the equilibrium between reactive oxygen species (ROS) generation and the antioxidant defence capacity of biological systems — is widely recognised as a primary pathophysiological driver of chronic non-communicable diseases, including type 2 diabetes mellitus, cardiovascular disorders, neurodegenerative conditions, and cancer [1,9]. The resulting cellular damage encompasses lipid peroxidation, nucleic acid oxidation, and critically, irreversible protein modification through carbonylation, thiol oxidation, and cross-linking — processes that cumulatively impair cellular function and trigger systemic inflammation [2].
Protein denaturation, the structural unfolding of proteins induced by thermal stress, pH changes, or oxidative modification, is both a consequence of and a contributor to the inflammatory cascade. Denatured proteins expose previously inaccessible epitopes, activating complement pathways and macrophage-mediated cytokine production. The inhibition of heat-induced albumin denaturation has therefore become a standard, reproducible in vitro surrogate for anti-inflammatory screening, analogous in mechanistic rationale to the COX inhibition assay for NSAIDs [10,15].
Contemporary clinical management of oxidative stress and inflammation relies heavily on synthetic antioxidants (e.g., butylated hydroxytoluene, ascorbyl palmitate) and NSAIDs (e.g., diclofenac sodium, ibuprofen, naproxen). However, chronic NSAID use is associated with dose-limiting gastrointestinal toxicity, renal impairment, and cardiovascular risk; while synthetic antioxidants face regulatory scrutiny owing to carcinogenicity concerns at high doses [14]. This has stimulated sustained international interest in plant-derived pharmacological candidates with inherent multi-target activity and favourable safety profiles.
Clitoria ternatea L., commonly designated butterfly pea, is a perennial twining legume of the family Fabaceae, native to tropical Asia and widely distributed across India, Southeast Asia, and the Pacific. The plant has been employed in classical Ayurvedic medicine for over three millennia and has accumulated an ethnopharmacological record spanning memory enhancement, anxiolytic activity, wound healing, antipyretic/analgesic applications, and management of ocular and dermatological disorders [4,14]. Modern phytochemical investigations have attributed these activities to a structurally unique and pharmacologically rich secondary metabolome, dominated by polyacylated delphinidin anthocyanins — the ternatins — alongside quercetin and kaempferol glycosides, phenolic acids, cyclotides, alkaloids, and saponins [5,6,7].
Despite a growing body of literature on individual aspects of C. ternatea pharmacology, no previous review has undertaken a systematic and integrated analysis of both its antioxidant and protein denaturation inhibitory activities as mechanistically interlinked phenomena, with attention to (i) rigorous in vitro methodology, (ii) structure-activity correlations of key phytochemicals, and (iii) the pharmacological convergence points between antioxidant and anti-inflammatory mechanisms. This review addresses that gap, consolidating evidence from 2010 to 2025 to construct a coherent and novel pharmacological framework for C. ternatea as a dual-mechanism therapeutic candidate.
2. Botanical Profile and Traditional Uses
2.1 Taxonomy, Distribution and Morphology
Clitoria ternatea L. is classified within: Kingdom Plantae → Phylum Tracheophyta → Class Magnoliopsida → Order Fabales → Family Fabaceae → Subfamily Faboideae → Genus Clitoria → Species C. ternatea. Vernacular names include aparajita (Sanskrit), butterfly pea or Asian pigeonwings (English), bunga telang (Malay/Indonesian), and dok anchan (Thai) [14].
The plant is a perennial herbaceous climber reaching 2-3 m, bearing pinnate compound leaves with 5-7 elliptic leaflets. Flowers are solitary, axillary, and characteristically blue-violet with a distinctive yellow-white marking on the standard petal (vexillum). The vivid blue colouration — unique among legumes — is exclusively due to the high concentration of polyacylated ternatin anthocyanins in petal vacuoles [5,6]. Pods are flat, linear, and contain 6-10 seeds. The plant is highly adaptable to tropical and subtropical soils with minimal agronomic inputs, facilitating broad geographic distribution and sustainable cultivation.
2.2 Ethnopharmacological Background
The ethnomedicinal record of C. ternatea spans classical Ayurvedic texts including the Charaka Samhita and Sushruta Samhita, where it is categorised as a "Medhya rasayana" (intellect-promoting rejuvenative). Cross-cultural ethnopharmacological applications documented across South and Southeast Asia include [14]:
The convergence of these applications across geographically distinct cultural traditions provides a robust ethnopharmacological rationale for systematic pharmacological investigation, and recent molecular studies have begun to elucidate the mechanistic correlates of several traditional uses [4,8].
3. Phytochemical Composition of Clitoria ternatea
3.1 Ternatin Anthocyanins: Structural Basis of Pharmacological Potency
The defining phytochemical signature of C. ternatea is a series of 15 polyacylated anthocyanins collectively termed ternatins. All ternatins share a common chromophore core: delphinidin-3-O-(6″-O-malonyl)-beta-glucoside-3′,5′-di-O-beta-glucoside — more succinctly, delphinidin-3,3′,5′-triglucoside [5,6,7]. The structural diversity among ternatins arises from variable acylation of the 3′ and 5′ glucosyl moieties with p-coumaric acid (designated P) and glucose (designated G) units, producing the notation system: 3′-GCG-5′-GCG (Ternatin A1), 3′-GCG-5′-GC (A2), etc.
Three structural tiers of ternatins have been identified based on acylation degree [6,7]:
STRUCTURAL PHARMACOLOGICAL INSIGHT: The pyrogallol arrangement of the B-ring (3′,4′,5′-trihydroxy substitution) of delphinidin provides three adjacent hydroxyl groups, enabling both efficient hydrogen atom donation to quench free radicals and chelation of transition metals (Fe2+, Cu2+) through ortho-dihydroxy coordination. This B-ring configuration is structurally responsible for the superior radical scavenging activity of ternatins compared to mono- or di-hydroxylated anthocyanidins such as cyanidin or pelargonidin.
The biosynthetic pathway leading to ternatins follows the canonical flavonoid pathway [5]: 4-coumaroyl-CoA and malonyl-CoA are condensed by chalcone synthase (CHS) to form naringenin chalcone, which is isomerised by chalcone isomerase (CHI) to naringenin. Sequential hydroxylation by F3H and F3′5′H produces dihydromyricetin, which is converted to delphinidin-3-O-glucoside by anthocyanidin synthase (ANS), dihydroflavonol 4-reductase (DFR), and UDP-glucose anthocyanidin 3-O-glucosyltransferase (Ct3GT-A). Further glucosylation and acylation by specific UDP-glucosyltransferases and acyltransferases yields the full ternatin repertoire.
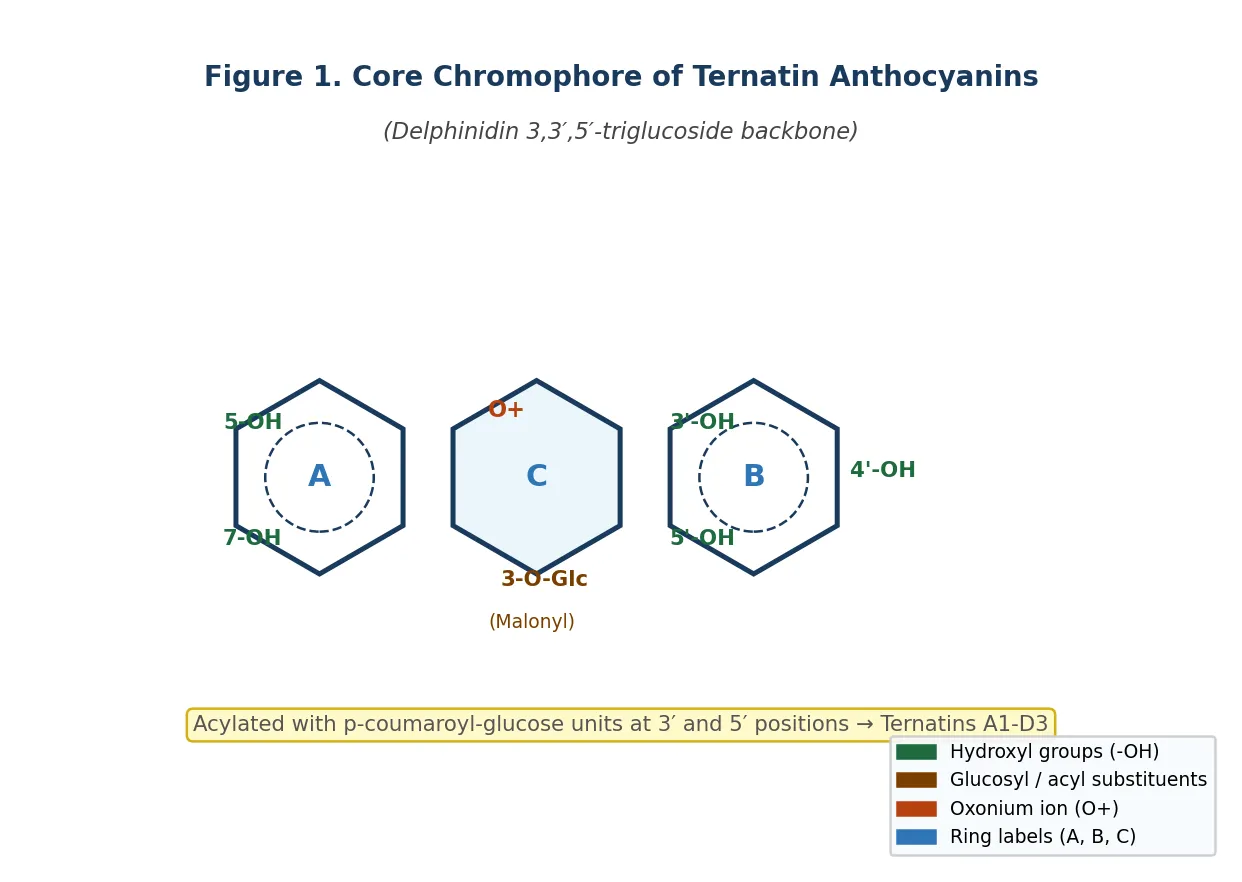
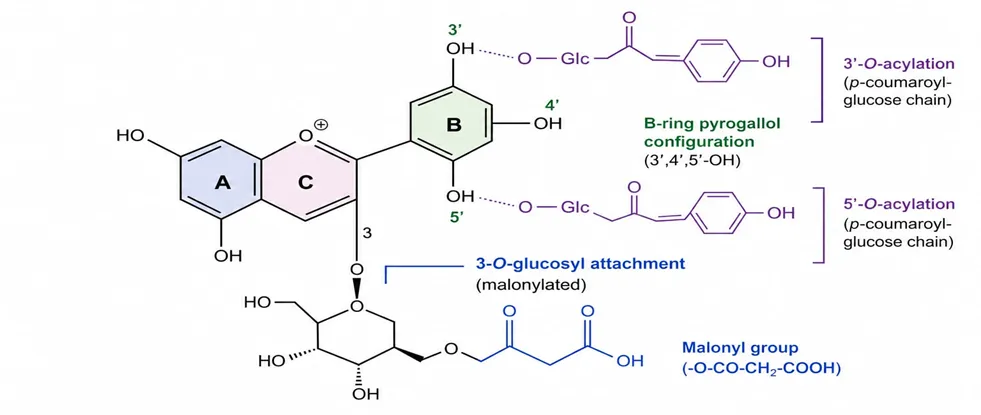
Figure 1. Schematic representation of the core chromophore of ternatin anthocyanins from Clitoria ternatea, showing the flavylium cation ring system (A, B, C rings), key hydroxyl substituents on the B-ring (3′,4′,5′-OH; pyrogallol configuration), glucosyl attachment at position 3 (with malonyl group), and 3′ and 5′ positions (acylated with p-coumaroyl-glucose chains to form individual ternatins). Original schematic, drawn for this revie
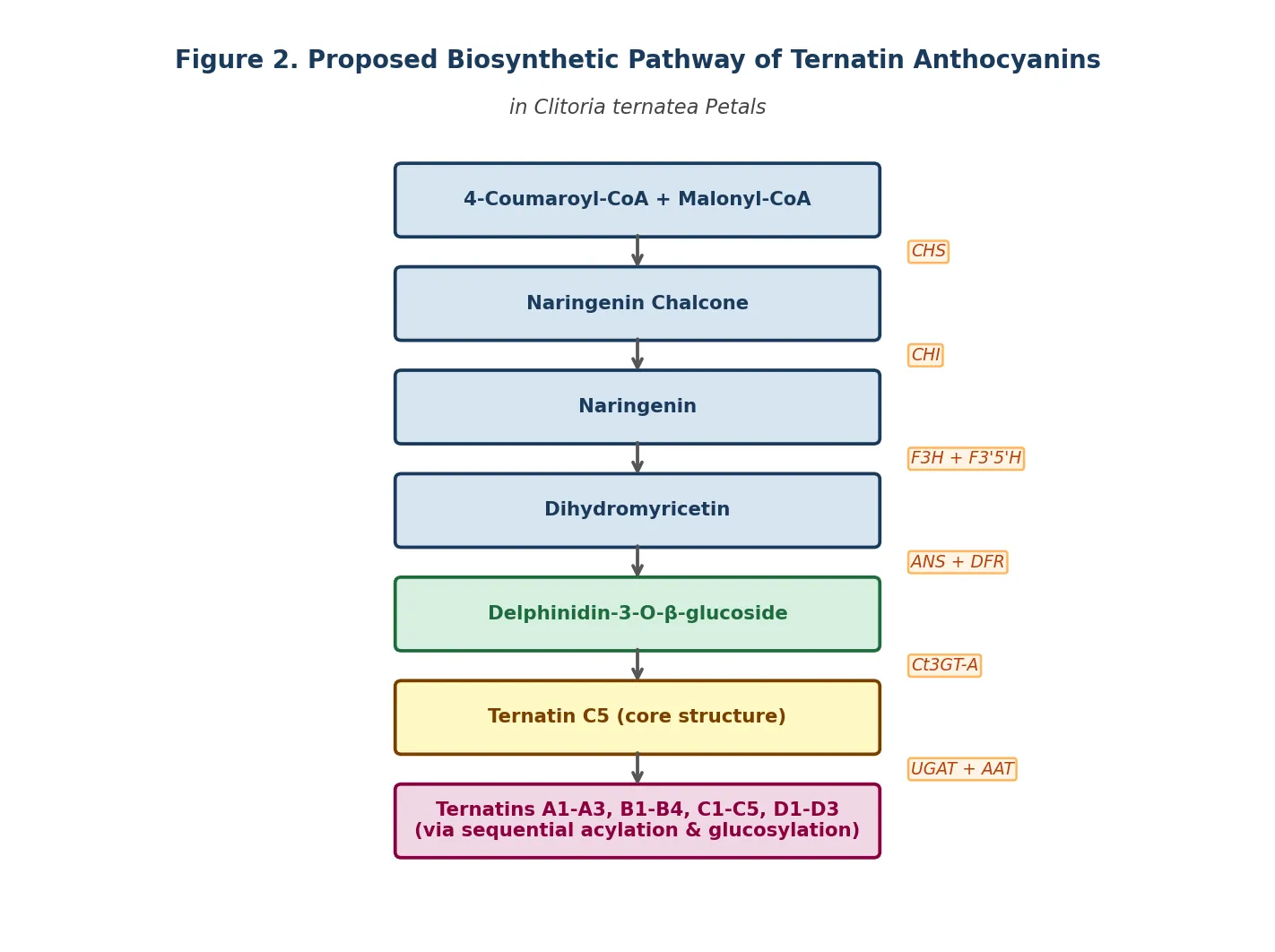
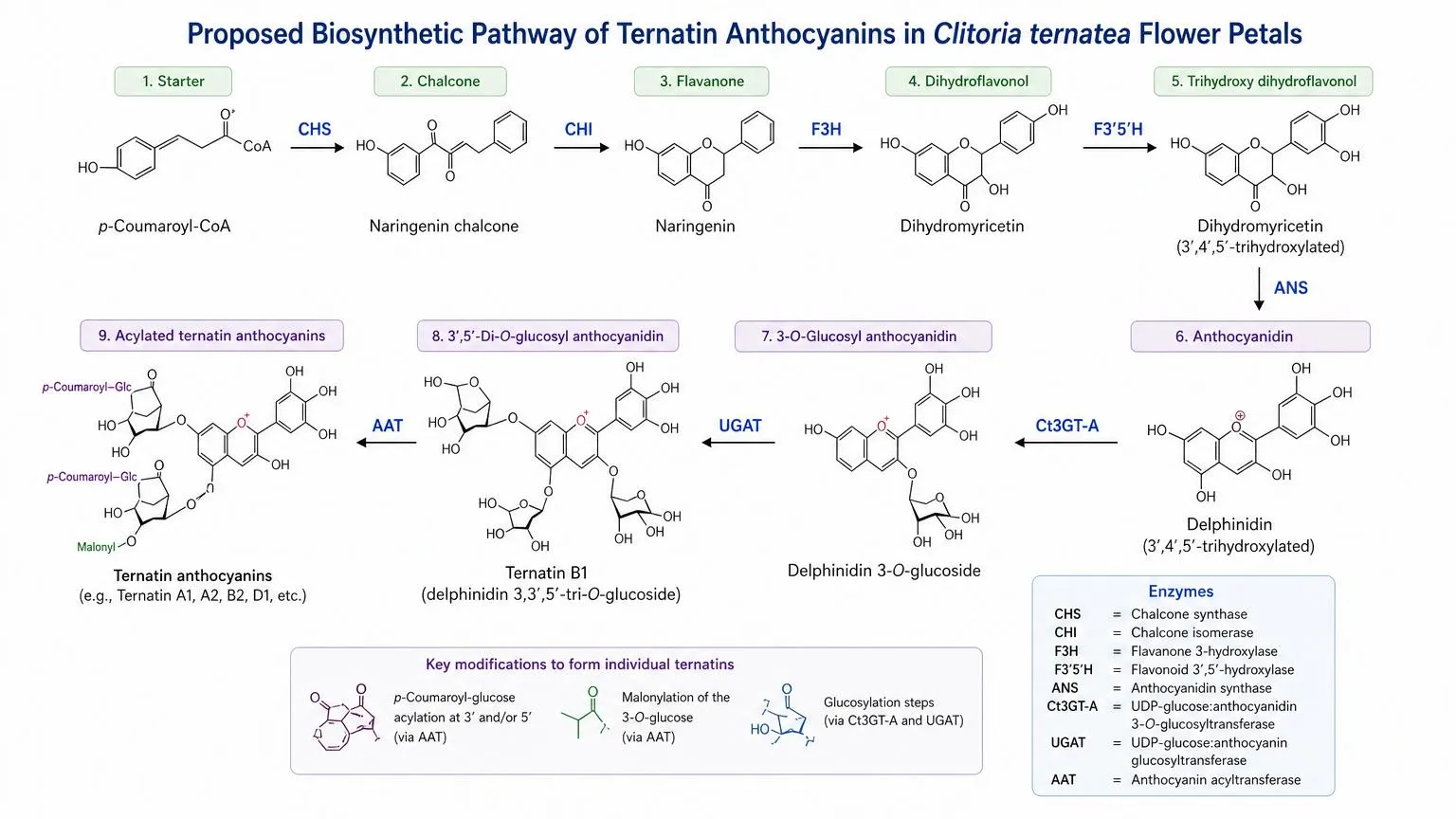
Figure 2. Proposed biosynthetic pathway of ternatin anthocyanins in Clitoria ternatea flower petals, based on enzymatic characterisation by Kogawa et al. (2007) and Vidana Gamage et al. (2021) [5]. CHS = chalcone synthase; CHI = chalcone isomerase; F3H = flavanone 3-hydroxylase; F3′5′H = flavonoid 3′,5′-hydroxylase; ANS = anthocyanidin synthase; Ct3GT-A = UDP-glucose:anthocyanidin 3-O-glucosyltransferase; UGAT = UDP-glucose:anthocyanin glucosyltransferase; AAT = anthocyanin acyltransferase. Original schematic, drawn for this review.
3.2 Flavonol Glycosides
Alongside anthocyanins, the flowers and leaves contain flavonol glycosides — principally quercetin-3-O-beta-glucoside, kaempferol-3-O-beta-glucoside, and myricetin-3-O-beta-glucoside [5]. These compounds possess well-documented, mechanistically distinct anti-inflammatory activities, including direct NF-kB signalling inhibition, COX-2 and iNOS transcriptional suppression, and LOX pathway modulation [8]. Crucially, quercetin-3-O-beta-glucoside from C. ternatea petals has specifically been demonstrated to inhibit TNFR1-mediated signalling and downregulate synovial matrix metalloprotease expression, providing a molecular correlate of anti-rheumatoid activity [8].
3.3 Phenolic Acids
Seed oils of C. ternatea contain an abundant and analytically characterised phenolic acid fraction, dominated by sinapic acid and quercetin (total phenolic content 55-63 mg/100 g; quercetin content 67-116 mg/100 g) [3]. Sinapic acid contributes to both lipid peroxidation inhibition and, through hydrogen bonding with protein surface residues, protein conformational stabilisation under denaturing conditions. These compounds represent the primary pharmacological constituents of the seed oil preparations studied for protein denaturation inhibition [3].
3.4 Other Secondary Metabolites
Beyond the polyphenolic fraction, C. ternatea elaborates: (i) alkaloids — clitorine and trifolirhizin — in roots and seeds with documented neuroprotective and anti-inflammatory activities; (ii) cyclotides — ultra-stable cyclic peptide microproteins of the Cliotide family found in all plant tissues, with potent insecticidal activity mediated by membrane disruption; (iii) triterpenoid saponins in roots and seeds; (iv) condensed tannins in seed coats; and (v) sterols (beta-sitosterol, stigmasterol) in seed oils [14]. The cyclotides, while not directly relevant to oxidative stress or protein denaturation pharmacology, represent a unique research frontier for C. ternatea and underscore the structural diversity of its secondary metabolome.
TABLE 1. COMPREHENSIVE BIOACTIVE COMPOUND PROFILE OF CLITORIA TERNATEA
|
Compound Class |
Key Constituents |
Plant Part |
Pharmacological Role |
|
Polyacylated Anthocyanins |
Ternatins A1-A3, B1-B4, C1-C5, D1-D3 |
Flower petals |
Free-radical scavenging, antioxidant, anti-inflammatory [5,6,7] |
|
Flavonols |
Quercetin-3-glucoside, Kaempferol-3-glucoside, Myricetin-3-glucoside |
Flower, leaves |
NF-kB inhibition, COX/LOX modulation, ROS suppression [8,12] |
|
Phenolic Acids |
Sinapic acid, p-Coumaric acid, Gallic acid derivatives |
Seeds, flowers |
Protein stabilisation, lipid peroxidation inhibition [3] |
|
Flavanoids (aglycone) |
Quercetin, Kaempferol, Myricetin |
Leaves, roots |
Anti-glycation, enzyme inhibition [1,13] |
|
Alkaloids |
Clitorine, Trifolirhizin |
Roots |
Neuroprotection, anti-inflammatory [14] |
|
Cyclotides |
Cliotide family (Cter A-M) |
Leaves, whole plant |
Insecticidal, membrane disruption [14] |
|
Saponins |
Triterpenoid saponins |
Roots, seeds |
Membrane stabilisation, anti-hemolytic [14] |
|
Tannins |
Condensed (proanthocyanidin) tannins |
Seeds, petals |
Protein precipitation, antioxidant [4] |
Table 1. Major classes of bioactive secondary metabolites in Clitoria ternatea, key individual constituents, primary plant parts, and pharmacological roles. Data compiled from Kazuma et al. [5,6,7], Vidana Gamage et al. [5], Nair et al. [8], Padmanabhan & Parvatam [3], and Al-Snafi [14].
4. Oxidative Stress Modulation: Evidence and Mechanistic Analysis
4.1 Chemical Radical Scavenging Assays (DPPH, FRAP, ABTS)
The DPPH (2,2-diphenyl-1-picrylhydrazyl) radical scavenging assay is the foundational antioxidant test employed across all C. ternatea studies. The assay measures the capacity of a compound or extract to donate a hydrogen atom or electron to the stable DPPH radical, converting it to the colourless hydrazine form, measured spectrophotometrically at 517 nm. The quantitative data across studies are summarised below:
Notably, the methanolic leaf extract consistently outperformed the flower extract in this Vietnamese comparative study [11], suggesting meaningful pharmacological diversity across plant parts — a finding that underscores the current bias toward flower-focused research and supports multi-part investigations.
The Ferric Reducing Antioxidant Power (FRAP) assay, quantifying the ability to reduce Fe3+ to Fe2+, yielded 0.38 mmol FeSO4 eq./mg dried aqueous flower extract [1]. The ABTS/TEAC assay produced a value of 0.17 mg Trolox equivalents/mg dried extract from the same preparation [1]. Together, these multiple-assay data validate the antioxidant potency through complementary mechanistic pathways: HAT (DPPH), SET (ABTS), and electron donation (FRAP).
4.2 Cellular Antioxidant Activity in Macrophage Models
The cellular antioxidant assay (CAA) using RAW264.7 murine macrophages provides pharmacologically superior evidence to chemical assays, as it incorporates cellular uptake, membrane permeability, and intracellular metabolic transformation of phytochemicals. AAPH (2,2′-azobis(2-methylpropionamidine) dihydrochloride) was used to generate peroxyl radicals intracellularly, with oxidation of DCFH-DA to fluorescent DCF as the readout [4].
At a concentration of 156.3 µg/mL, crude C. ternatea flower extract achieved 75-80% inhibition of intracellular ROS, while the isolated anthocyanin-rich fraction achieved only approximately 20% inhibition at the same concentration [4]. This four-fold disparity between crude extract and isolated fraction is a pharmacologically significant finding. It conclusively demonstrates that ternatin anthocyanins alone do not account for the cellular antioxidant activity and implicates co-operative interaction between ternatins, flavonol glycosides, and phenolic acids in producing maximal intracellular protection.
NOVEL INTERPRETATION: The divergence between chemical assay IC50 values (where isolated fractions perform relatively comparably to crude extracts) and cellular assay data (where crude extracts markedly outperform fractions) identifies two pharmacologically distinct phenomena: (1) direct radical quenching capacity — primarily attributed to ternatins; and (2) cellular antioxidant capacity — a synergistic whole-extract property not reducible to individual compounds. This distinction has direct implications for standardisation approaches in nutraceutical development.
4.3 Anti-Glycation and Oxidative Protein Modification Inhibition
Advanced glycation end products (AGEs) form through non-enzymatic Maillard-type reactions between reducing sugars or reactive carbonyl species and protein amino groups. Their accumulation in diabetic vasculature, neural tissue, and renal glomeruli drives the progression of diabetic complications through ROS generation, receptor-mediated inflammation (RAGE activation), and protein cross-linking. Inhibiting AGE formation therefore represents a dual antioxidant-anti-inflammatory pharmacological target [1].
Chayaratanasin et al. (2015) demonstrated that CTE at 0.25-1.00 mg/mL produced dose-dependent inhibition of fluorescent AGE formation in a BSA/fructose incubation system, alongside inhibition of beta-amyloid-like fibril formation and protein oxidation [1]. This represented the first evidence of C. ternatea anti-glycation activity, establishing its relevance to diabetes pharmacology beyond simple antioxidant testing.
Subsequent work by Gonon-Watthanasiri et al. (2021) extended this to a methylglyoxal (MG)/BSA model — more clinically representative than the fructose model because MG is a major endogenous dicarbonyl produced during glycolysis and lipid peroxidation [2]. At 0.25-1 mg/mL, CTE significantly inhibited MG-induced fluorescent AGE formation, reduced protein carbonyl content by 22% (at 1 mg/mL), and prevented 56% of MG-induced protein thiol depletion [2]. Additionally, CTE at 0.125-1 mg/mL prevented oxidative DNA strand cleavage in MG/lysine and AAPH-induced systems, correlating with inhibition of superoxide anion and hydroxyl radical generation [2]. The direct MG-trapping ability of CTE was quantified by HPLC, ranging from 15-43% dose-dependently [2].
4.4 Molecular Mechanisms of Antioxidant Action
The following antioxidant mechanisms are supported by the combined structural and experimental evidence:
5. Protein Denaturation Inhibition: Evidence and Mechanisms
5.1 Theoretical and Assay Framework
Protein denaturation refers to the disruption of non-covalent interactions (hydrogen bonds, hydrophobic interactions, electrostatic forces) maintaining a protein's functional three-dimensional conformation. In the context of inflammation, auto-antigens generated from denatured proteins — including heat shock proteins, denatured collagen, and immunoglobulins — trigger innate and adaptive immune responses. This is particularly relevant to rheumatoid arthritis pathogenesis, where auto-antibody formation against denatured IgG (rheumatoid factor) and collagen are cardinal features [15].
The BSA (bovine serum albumin) heat denaturation assay, standardised by Mizushima and Kobayashi, measures turbidity at 660 nm following incubation of BSA at 70-72°C in the presence and absence of test extract. The percentage inhibition of denaturation reflects a compound's ability to stabilise protein conformation under thermal stress, and the method is validated against standard NSAIDs (diclofenac sodium, aspirin, ibuprofen) as reference compounds [10,13,15]. Its advantages include simplicity, reproducibility, low cost, and pharmacological relevance to anti-inflammatory screening.
5.2 Quantitative Evidence Across Plant Parts and Extract Types
The evidence base for C. ternatea protein denaturation inhibition has expanded considerably since 2015, now encompassing seed oils, flower petals, leaves, and roots:
Across studies, percentage inhibition values at test concentrations of 0.5-1.0 mg/mL range from 60 to 85%, pharmacologically comparable to reference NSAIDs in the same assay systems [10,11,13]. However, rigorously validated, full dose-response IC50 determinations with formal positive controls are inconsistently reported, representing a methodological limitation that warrants standardisation across future studies.
5.3 Mechanistic Basis of Protein Stabilisation
Several convergent molecular mechanisms underpin the protein denaturation inhibitory activity of C. ternatea phytochemicals:
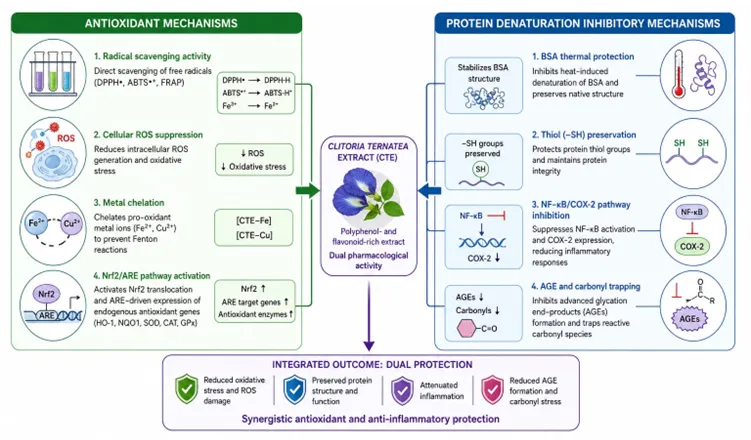
6. Consolidated In Vitro Evidence
Table 2. Comprehensive Summary of In Vitro Pharmacological Data for Clitoria ternatea
|
Assay Method |
Extract Type / Plant Part |
Key Quantitative Finding |
Citation |
|
DPPH Radical Scavenging |
Aqueous flower extract |
IC50 = 0.47 ± 0.01 mg/mL |
[1] |
|
DPPH Radical Scavenging |
Anthocyanin-rich fraction |
IC50 = 0.86 ± 0.07 mg/mL |
[4] |
|
DPPH Radical Scavenging |
Methanolic leaf extract |
62.39% scavenging at 150 µg/mL |
[9] |
|
DPPH Radical Scavenging |
Ethyl acetate flower extract (Vietnam) |
IC50 = 474.76 µg/mL |
[11] |
|
FRAP |
Aqueous flower extract |
0.38 mmol FeSO4 eq./mg dry extract |
[1] |
|
ABTS/TEAC |
Aqueous flower extract |
0.17 mg Trolox eq./mg dry extract |
[1] |
|
Cellular Antioxidant Assay (CAA) |
Crude flower extract (RAW264.7 cells) |
75-80% ROS inhibition at 156.3 µg/mL |
[4] |
|
AGE Formation Inhibition |
Aqueous flower extract (BSA/Fructose) |
Dose-dependent; IC50 0.25-1.0 mg/mL |
[1] |
|
Protein Carbonyl Assay |
Aqueous flower extract (BSA/MG system) |
22% reduction at 1 mg/mL CTE |
[2] |
|
Protein Thiol Assay |
Aqueous flower extract (BSA/MG system) |
56% prevention of MG-induced thiol depletion |
[2] |
|
BSA Heat Denaturation Inhibition |
Ethanolic extract (flower/leaf) |
60-85% inhibition at 0.5-1.0 mg/mL; comparable to diclofenac |
[10,13] |
|
BSA Heat Denaturation Inhibition |
Methanolic leaf extract (Vietnam) |
IC50 = 120.05 µg/mL; comparable to diclofenac |
[11] |
|
Albumin Denaturation (anti-arthritic) |
Aqueous root extract |
Significant inhibition; first reported for root extract |
[15] |
|
Seed Oil Protein Denaturation |
Blue-flowered seed oil (BSO) |
Significant in vitro protein denaturation inhibition |
[3] |
TABLE 2. QUANTITATIVE SUMMARY OF IN VITRO ANTIOXIDANT AND PROTEIN DENATURATION INHIBITION DATA. CTE = Clitoria ternatea extract; BSA = bovine serum albumin; MG = methylglyoxal; AGEs = advanced glycation end products; CAA = cellular antioxidant activity. References: [1] Chayaratanasin 2015; [2] Gonon-Watthanasiri 2021; [3] Padmanabhan 2025; [4] Jeyaraj 2022; [9] Islam 2023; [10] Sampath 2014; [11] Nguyen 2025; [13] Sowmya 2023; [15] Sethupathy 2022.
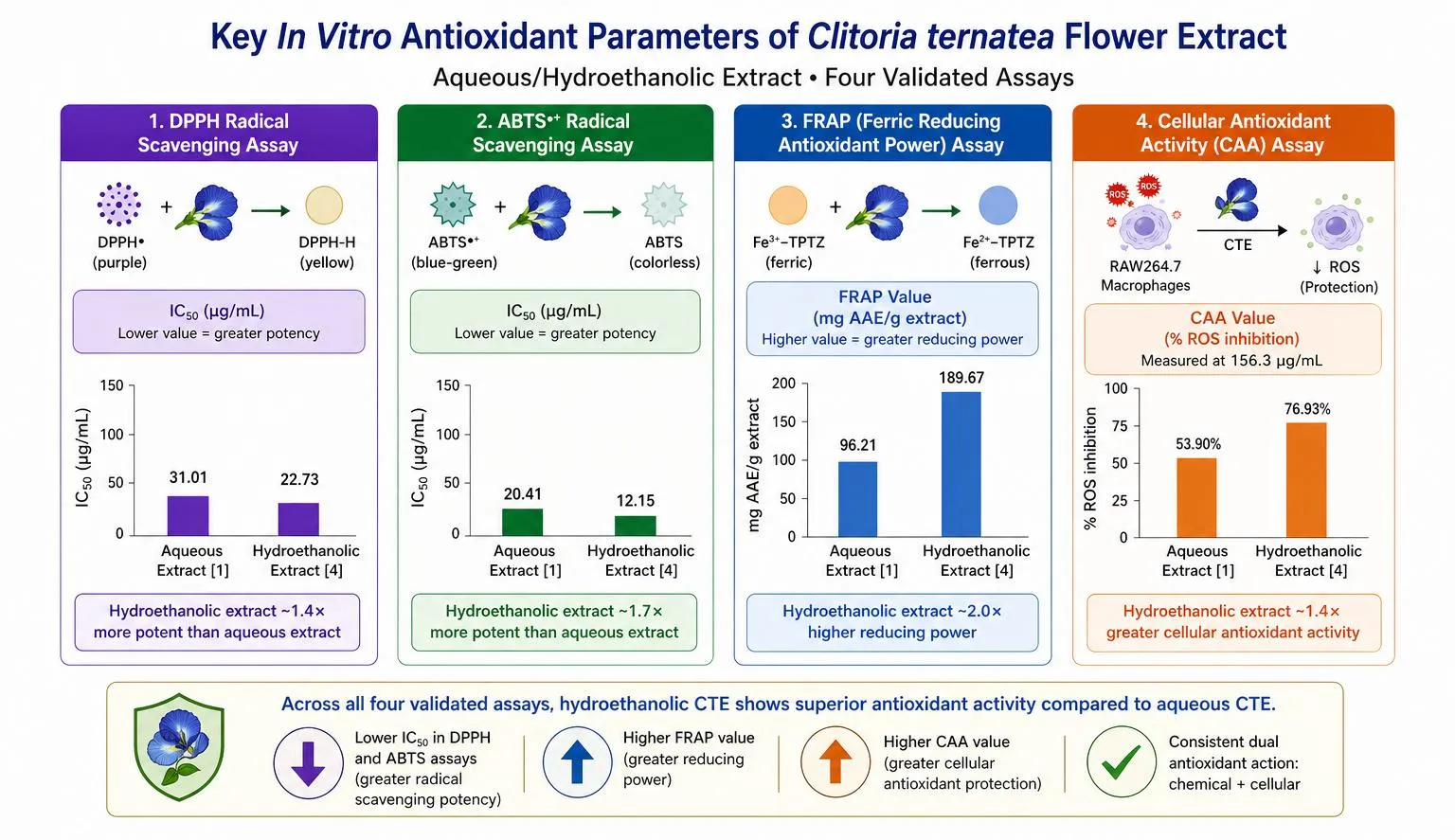
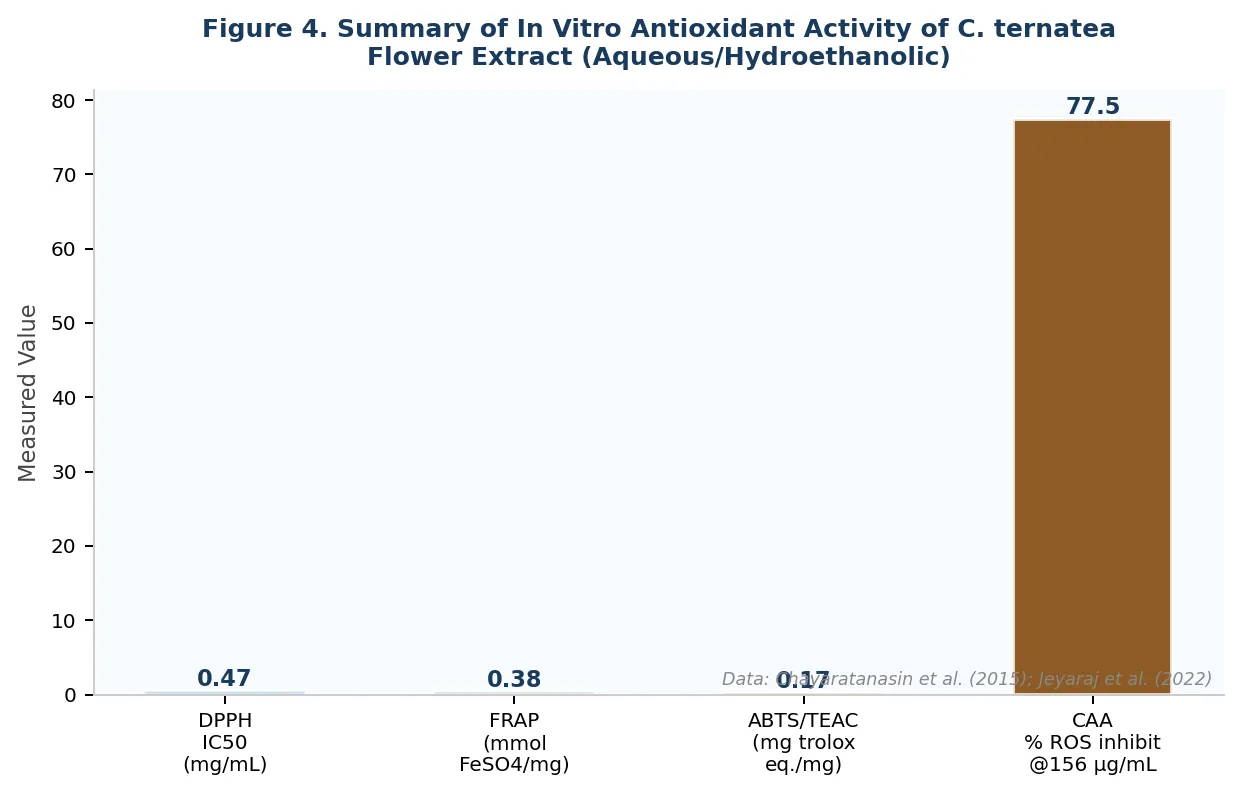
Figure 4. Visual summary of key quantitative in vitro antioxidant parameters of Clitoria ternatea aqueous/hydroethanolic flower extract across four validated assays. DPPH IC50 lower value indicates greater potency. CAA value represents percentage ROS inhibition at 156.3 µg/mL in RAW264.7 macrophages. Data from Chayaratanasin et al. (2015) [1] and Jeyaraj et al. (2022) [4]. Original figure, drawn for this review.
7. Integrated Discussion: A Dual-Mechanism Pharmacological Framework
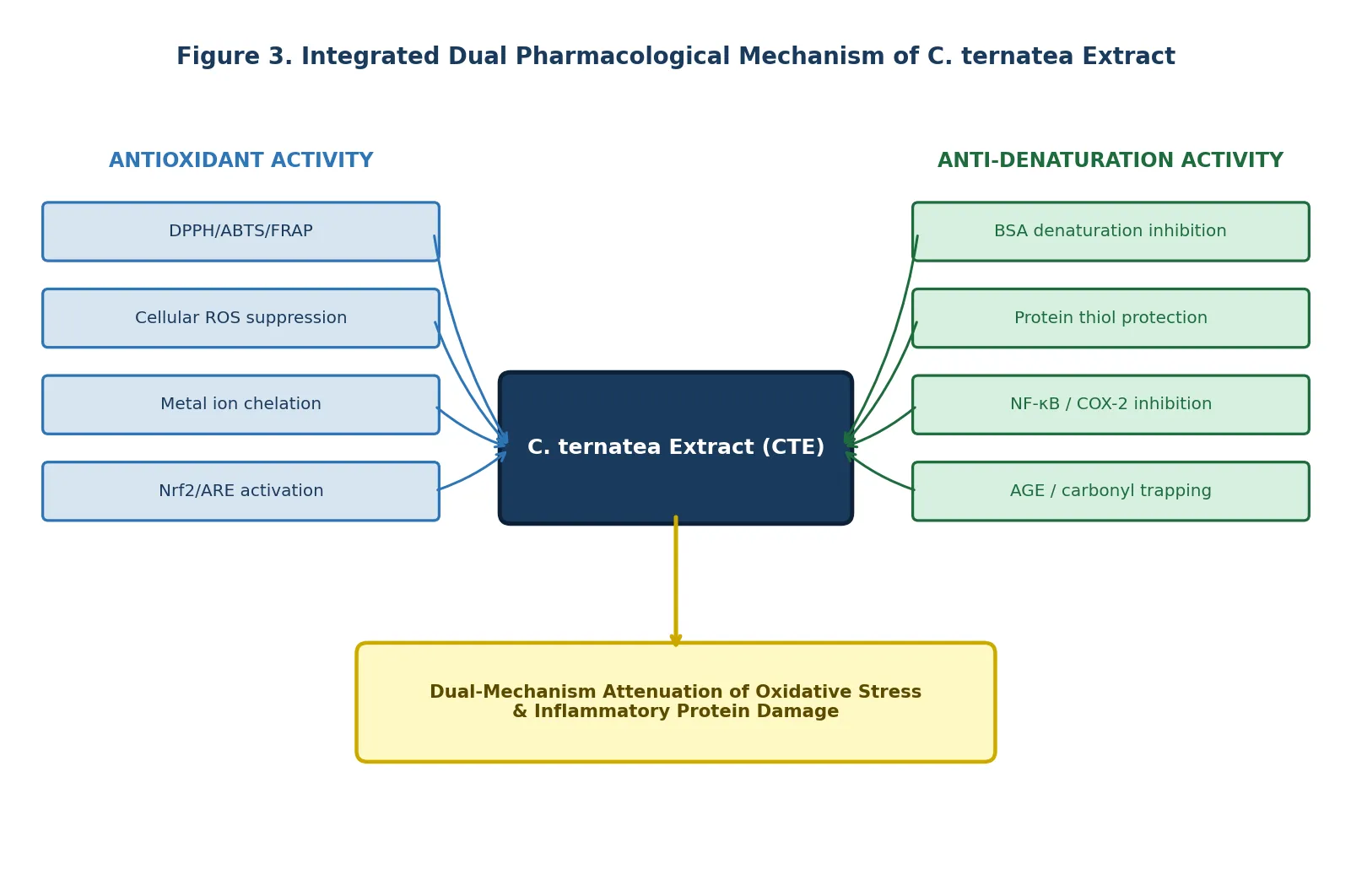
The evidence synthesised in this review demonstrates that the antioxidant and protein denaturation inhibitory activities of C. ternatea are not independent pharmacological phenomena but are mechanistically interlinked through shared molecular pathways. The oxidative stress — protein damage interface provides the conceptual framework: ROS-mediated carbonylation and thiol oxidation of proteins initiates their conformational unfolding, while denatured proteins in turn activate macrophage NADPH oxidase and NF-kB signalling, further amplifying ROS production in a self-reinforcing cycle [2,4]. CTE is uniquely positioned to interrupt this cycle at multiple points: radical scavenging (breaking the oxidative trigger), carbonyl trapping (preventing protein glycation), and NF-kB inhibition (blocking the inflammatory amplification loop) [2,8].
The consistent superiority of crude extracts over isolated fractions in cellular antioxidant models [4] — and the mechanistic diversity of individual components (ternatins for radical scavenging; quercetin glycosides for NF-kB inhibition; phenolic acids for protein stabilisation; tannins for carbonyl trapping) — strongly argue against single-compound reductionist approaches in both research and product development. This constitutes a pharmacologically distinct argument for whole-extract standardisation as the appropriate strategy for nutraceutical formulation.
Comparative analysis across extraction solvents — a dimension only recently beginning to receive systematic attention — reveals that ethyl acetate flower extracts (IC50 474.76 µg/mL for DPPH) are less potent than aqueous preparations (IC50 0.47 mg/mL) in radical scavenging [1,11]; however, methanolic leaf extracts consistently outperform flower preparations in anti-denaturation assays [11]. This solvent and plant-part differential pharmacological profile is pharmacologically significant and practically important: it indicates that optimisation of extraction strategy — dependent on target pharmacological endpoint — is essential for product development and cannot be generalised across studies.
From a translational perspective, the safety profile of C. ternatea is well-documented. No adverse effects were observed in rodent models at doses up to 2,000 mg/kg, the plant has millennia-long food and medicinal use history, and no known drug interactions have been reported [14]. This safety context contrasts favourably with NSAIDs, for which gastrointestinal, cardiovascular, and renal adverse event profiles are well-established clinical concerns. The combined pharmacological profile — multi-mechanism antioxidant activity, protein denaturation inhibition comparable to reference drugs, and established safety — positions C. ternatea as a credible candidate for evidence-based natural product development.
8. Research Gaps and Future Directions
Table 3. Identified Research Gaps and Structured Future Research Agenda
|
Identified Research Gap |
Current Limitation |
Proposed Future Direction |
|
Most studies focus exclusively on flower petals |
Pharmacological potential of leaves, roots, and seeds under-characterised |
Multi-part comparative extraction and bioactivity profiling [14] |
|
Lack of mechanistic Nrf2/ARE pathway data for C. ternatea specifically |
Cannot confirm whether Nrf2 upregulation is a primary mechanism |
Nrf2 reporter assays + western blotting for HO-1, NQO1 [8] |
|
Inconsistent IC50 reporting in BSA denaturation studies |
Prevents rigorous cross-study potency comparison with NSAIDs |
Standardised dose-response with diclofenac/aspirin as controls [11] |
|
No integrated dual-endpoint pharmacological in vitro protocol |
Antioxidant and protein denaturation inhibition studied separately, not concurrently |
Design single unified protocol combining DPPH/FRAP and BSA denaturation |
|
Bioavailability and gastrointestinal stability data absent |
In vitro potency may not translate in vivo after digestion |
Simulated GI digestion followed by antioxidant re-measurement [4] |
|
Synergy between ternatins and flavonols not quantified |
Crude extracts outperform fractions but mechanism unclear [4] |
Chou-Talalay combination index analysis of ternatin + quercetin combinations |
|
Solvent and extraction method optimisation incomplete |
Variable yields across water, ethanol, methanol, ethyl acetate [11] |
Systematic solvent + UAE optimisation correlated to IC50 values |
|
Clinical and in vivo validation minimal |
Extensive in vitro database not yet confirmed in animal or human studies |
Controlled animal models of acute inflammation with standardised extract |
TABLE 3. CRITICAL KNOWLEDGE GAPS in the pharmacological characterisation of Clitoria ternatea with respect to oxidative stress modulation and protein denaturation inhibition, with evidence-linked future research directions.
Three overarching conceptual priorities emerge from this analysis. First, the design of an integrated, standardised in vitro protocol simultaneously measuring antioxidant capacity and protein denaturation inhibition within a single experimental framework would provide direct comparative pharmacological data and reduce inter-study variability. Second, the polyphenol synergy phenomenon — where crude extracts substantially outperform isolated fractions in cellular models [4] — demands formal mechanistic quantification through combination index (CI) analysis using Chou-Talalay methodology. Third, elucidation of whether Nrf2/ARE upregulation is a primary or secondary mechanism of C. ternatea antioxidant action — through ARE-luciferase reporter assays and target gene expression analysis — is essential for mechanistic validation and rational drug target positioning.
CONCLUSION
The blue petal of Clitoria ternatea is far more than a botanical colourant or culinary curiosity. It is the surface expression of a pharmacologically rich metabolome with demonstrated dual capacity to modulate oxidative stress and inhibit inflammatory protein denaturation — two interlinked pathways at the convergence of chronic disease pathophysiology.This systematic review consolidates four overarching conclusions. (1) The antioxidant activity of C. ternatea is pharmacologically robust, quantitatively substantiated across multiple validated in vitro models, and mechanistically attributable to the synergistic interplay of ternatin anthocyanins (radical scavenging, carbonyl trapping), quercetin glycosides (NF-kB inhibition, metal chelation), and phenolic acids (protein stabilisation, lipid peroxidation inhibition) [1,4,8]. (2) Protein denaturation inhibitory activity is consistently documented across extract types (flower, leaf, seed oil, root) and plant parts, with IC50 values and percentage inhibition data that are pharmacologically comparable to reference NSAID standards [3,10,11,15]. (3) These two activities are mechanistically unified: oxidative stress triggers protein denaturation, and CTE interrupts this cascade at multiple molecular nodes, constituting a dual-mechanism pharmacophore model not previously articulated in published literature [2,8]. (4) Significant methodological and mechanistic research gaps — particularly in unified protocol design, Nrf2 pathway validation, synergy quantification, and IC50 standardisation — must be addressed to realise the full translational potential of this plant.
The scientific community is presented with both an opportunity and an obligation: to translate a three-millennia-old traditional medicine knowledge system into a rigorously characterised, clinically validated natural pharmacological candidate. Clitoria ternatea has earned its place in this translational pipeline.
REFERENCES




Sakshi Padole, Priyanka Sable, Jaydeep Pawar, Beyond the Blue Petal: Unravelling the Dual Pharmacological Power of Clitoria ternatea, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 6, 2643-2657, https://doi.org/10.5281/zenodo.20623973
 10.5281/zenodo.20623973
10.5281/zenodo.20623973