
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
1College of Pharmaceutical Sciences, Pravara Institute of Medical Sciences (DU), Loni
Genome editing has progressed from meganucleases, zinc finger nucleases, and TALENs to CRISPR-Cas9, a bacterial/archaeal adaptive immune-derived platform that has democratized precise genetic manipulation through a programmable single guide RNA (sgRNA) directing Cas9 to DNA sequences adjacent to a protospacer adjacent motif (PAM; 5?-NGG-3? for SpCas9). This comprehensive review aims to integrate mechanistic foundations, cross-sector applications, and ethical/societal implications of CRISPR-Cas9 within a single critically evaluative framework. Drawing on a broad, interdisciplinary synthesis of published evidence, we describe key milestones from the first observation of CRISPR loci (1987) to mechanistic elucidation and sgRNA engineering (2012), rapid adoption for mammalian genome editing (2013), and expansion to >50,000 publications by 2023, alongside translation to approved therapy (FDA approval of Casgevy/exa-cel in 2023). Mechanistically, Cas9–sgRNA target recognition via PAM scanning, seed-dependent R-loop formation, and HNH/RuvC-mediated cleavage produces double-strand breaks repaired mainly by error-prone NHEJ/MMEJ (enabling knockouts) or less efficient, cell-cycle-restricted HDR (enabling precise insertions and point corrections). Major limitations include off-target editing, large deletions/rearrangements, p53-mediated DNA damage responses, delivery constraints (SpCas9 size vs AAV packaging), and immunogenicity from pre-existing anti-Cas9 immunity; mitigation strategies include GUIDE-seq/CIRCLE-seq/DISCOVER-seq profiling, high-fidelity Cas9 variants, paired nickases, anti-CRISPR proteins, and nanotechnology-enabled delivery (notably lipid nanoparticles and engineered extracellular vesicles). Applications span agriculture (disease resistance, yield and nutritional enhancement, edited livestock), medicine and clinical sciences (functional genomics screens, iPSC disease models, HbF reactivation for hemoglobinopathies, edited T cells, in vivo PCSK9 targeting), environmental science (gene drives, conservation genomics, monitoring), and biotechnology/bioinformatics (sgRNA design, off-target prediction, outcome quantification, metabolic engineering). Ethical analysis highlights germline editing risks underscored by the 2018 CCR5 embryo case, therapy–enhancement boundaries, equity and intellectual property concerns, and the need for adaptive, internationally coordinated governance reaffirmed in 2023. The review concludes that CRISPR-Cas9 is transforming precision biology, but responsible progress requires sustained technical refinement, rigorous safety evaluation, and inclusive governance to realize benefits for health, food security, and environmental sustainability.
The ability to precisely and efficiently alter the genetic composition of living organisms has long been a central ambition of molecular biology and biotechnology. Over the past four decades, researchers have developed a succession of genome-editing platforms — from meganucleases and zinc finger nucleases (ZFNs) to transcription activator-like effector nucleases (TALENs) — each representing a significant advance but also carrying limitations in terms of design complexity, cost, efficiency, or specificity [1,2]. The advent of CRISPR-Cas9 technology has fundamentally transformed this landscape, offering an unprecedented combination of simplicity, affordability, efficiency, and versatility that has propelled genome editing from specialist research laboratories into virtually every area of the life sciences [3,4].CRISPR-Cas9 is a programmable endonuclease system derived from an adaptive immune mechanism found in bacteria and archaea. The system employs a short synthetic single guide RNA (sgRNA) molecule to direct the Cas9 protein to a specific complementary DNA sequence in the genome, where it introduces a precise double-stranded break (DSB). The cell's own DNA repair machinery subsequently resolves the break — via non-homologous end joining (NHEJ) or homology-directed repair (HDR) — enabling the introduction of gene knockouts, point mutations, or targeted sequence insertions with high efficiency [5,6]. The relative ease with which the sgRNA can be redesigned to target virtually any genomic locus has made CRISPR-Cas9 accessible to researchers across disciplines, fundamentally democratizing genetic research [7].The publication of foundational studies by Doudna and Charpentier [8] and their simultaneous application to mammalian cell genomes by multiple research groups in 2013 triggered an explosion of scientific activity. Within a decade, CRISPR-Cas9 has been applied across agriculture, biomedical research, environmental science, clinical medicine, bioinformatics, and nanotechnology, and has catalysed the development of a new generation of CRISPR-based tools including base editors, prime editors, and CRISPR-based diagnostic platforms [9,10]. This unprecedented pace of development has been matched, however, by growing scrutiny of the technology's safety profile and the ethical implications of its potential applications — particularly those involving heritable changes to the human germline [11].Several comprehensive reviews have addressed individual dimensions of CRISPR-Cas9 technology, focusing respectively on its molecular mechanism [12], its clinical translation [13], or its applications in specific sectors such as agriculture [14] or oncology [15]. However, a unified, cross-disciplinary synthesis that integrates the mechanistic, applied, and ethical aspects of CRISPR-Cas9 within a single framework remains a critical need, particularly given the rapid evolution of the field. The present review aims to fulfil this need by providing a comprehensive, current, and critically evaluative overview of CRISPR-Cas9 technology.Specifically, this review: (i) describes the molecular architecture and mechanism of action of the CRISPR-Cas9 system; (ii) traces the history of its discovery and key milestones in its development; (iii) examines in depth its applications across agriculture, medicine, clinical sciences, environmental science, nanotechnology, bioinformatics, and biotechnology; (iv) surveys the ethical and societal controversies surrounding its use; and (v) outlines future directions and emerging next-generation tools. By integrating these dimensions, the review aims to provide researchers, clinicians, policymakers, and students with a rigorous and accessible foundation for understanding one of the most consequential scientific developments of the twenty-first century.
2. Background and Literature Review
2.1 Pre-CRISPR Genome Editing Technologies
The history of targeted genome editing predates CRISPR-Cas9 by several decades. The earliest programmable genome editing tools — meganucleases, or homing endonucleases — were identified in the 1990s as naturally occurring enzymes capable of recognizing and cleaving long DNA sequences of 14–40 base pairs [16]. Although useful in research contexts, meganucleases were difficult to engineer for new target sequences, limiting their practical utility. Zinc finger nucleases (ZFNs), developed in the late 1990s and early 2000s, represented the first widely used programmable genome editing platform. ZFNs consist of a zinc finger DNA-binding domain fused to the FokI endonuclease cleavage domain, and can be designed to target specific sequences by engineering the zinc finger modules [17]. However, ZFN design is technically challenging and expensive, and the technology is associated with significant off-target cleavage and cytotoxicity.Transcription activator-like effector nucleases (TALENs), introduced around 2010, offered improved targeting flexibility over ZFNs by exploiting modular DNA-binding domains from TAL effector proteins of plant pathogenic bacteria [18]. While TALENs demonstrated superior specificity and lower cytotoxicity than ZFNs, their large size and the labour-intensive process of assembling the repetitive TALE arrays remained significant obstacles to widespread adoption. The emergence of CRISPR-Cas9 in 2012–2013 overcame both the technical complexity of ZFNs and TALENs and the scalability limitations of earlier tools, establishing a new paradigm for genome editing [5,19].
2.2 Discovery and Evolution of the CRISPR System
The origins of CRISPR lie not in biotechnology but in microbiology. Repetitive DNA sequences interspersed with unique spacer sequences — now recognized as the hallmark of CRISPR loci — were first observed in the Escherichia coli genome by Ishino et al. in 1987 [20], though their biological significance was not understood at the time. Subsequent studies in the early 2000s established that CRISPR sequences were found across a wide range of bacteria and archaea, and that the spacer sequences matched sequences from viruses (bacteriophages) and other mobile genetic elements [21]. This led to the hypothesis, confirmed experimentally by Barrangou et al. in 2007, that CRISPR arrays constitute an adaptive immune system enabling bacteria to resist viral infection by incorporating fragments of viral DNA as spacers [22].A series of landmark studies in the following years elucidated the mechanistic basis of CRISPR immunity. Brouns et al. demonstrated that CRISPR RNA (crRNA) — transcribed from the CRISPR array — guided Cas proteins to complementary sequences in invading viral DNA [23]. Gasiunas et al. and, in a parallel study, Jinek et al. showed that the Cas9 protein from Streptococcus pyogenes (SpCas9) was a dual RNA-guided endonuclease capable of cleaving both strands of target DNA [24,25]. Crucially, Jinek et al. demonstrated that the two RNA components required for Cas9 activity — crRNA and trans-activating crRNA (tracrRNA) — could be fused into a single chimeric sgRNA without loss of function, simplifying the system considerably and laying the groundwork for its application as a programmable genome-editing tool [25].The landmark 2012 paper by Doudna and Charpentier in Science described the biochemistry of the SpCas9–sgRNA system in detail and demonstrated its utility for targeted cleavage of purified DNA in vitro [8]. Within months, three simultaneous publications in 2013 — from the laboratories of Zhang, Church, and Bhatt — demonstrated efficient CRISPR-Cas9-mediated genome editing in human and mouse cells, establishing the technology's clinical and research potential [26,27]. Since these foundational studies, the field has advanced with extraordinary speed. By 2023, more than 50,000 publications on CRISPR-Cas9 had appeared in the scientific literature, and the technology was being applied across virtually every domain of biology and medicine [9].
3. CRISPR-Cas9 Technology: Molecular Mechanism and Components
3.1 Molecular Architecture of the CRISPR-Cas9 System
The CRISPR-Cas9 system as adapted for genome editing consists of two primary components: the Cas9 endonuclease protein and the single guide RNA (sgRNA). The Cas9 protein from Streptococcus pyogenes (SpCas9) — the most widely used variant — is a large (approximately 160 kDa) multi-domain enzyme containing two nuclease lobes and a recognition (REC) lobe [28]. The two nuclease domains, HNH and RuvC, cleave the strand complementary to the sgRNA and the non-complementary strand, respectively, resulting in blunt-ended double-stranded DNA breaks [29]. The REC lobe mediates interaction with the sgRNA and is responsible for target recognition and conformational activation of the nuclease domains upon DNA binding (Figure 1).
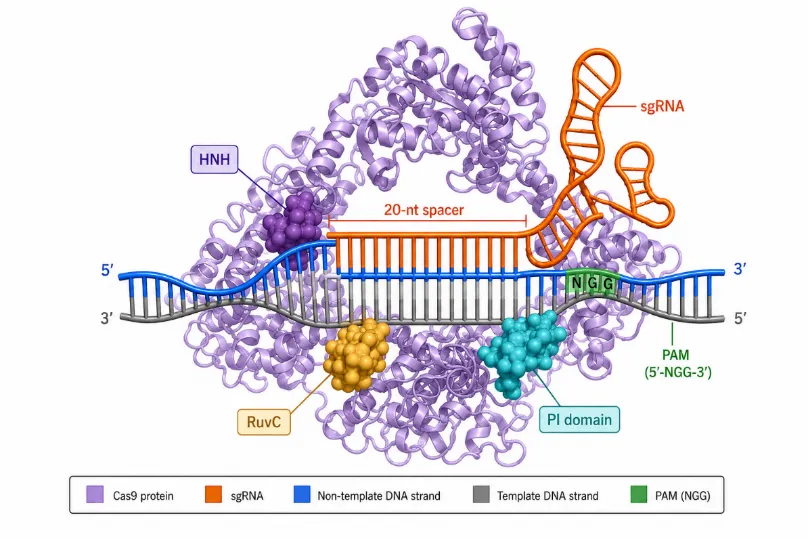
Figure 1: Molecular architecture of the CRISPR-Cas9 complex.
The Cas9 protein (shown in ribbon representation) is depicted in complex with the sgRNA (orange) and a target DNA duplex (blue: non-template strand; grey: template strand). Key functional domains — HNH, RuvC, and the PAM-interacting (PI) domain — are labelled. The 20-nt spacer sequence in the sgRNA is complementary to the protospacer in the target DNA, immediately 5' of the PAM sequence (5'-NGG-3' for SpCas9).
The sgRNA is a chimeric ~100-nucleotide RNA molecule that combines the functions of the crRNA and tracrRNA [25]. Its 5' end contains a programmable 20-nucleotide spacer sequence complementary to the target DNA protospacer, while its 3' end forms a stem-loop scaffold that mediates binding to Cas9. A critical requirement for Cas9 activity is the presence of a short protospacer adjacent motif (PAM) immediately downstream of the protospacer on the non-template strand. For SpCas9, the canonical PAM sequence is 5'-NGG-3', which constrains the choice of targetable sequences to those adjacent to an NGG dinucleotide [30]. Alternative Cas9 orthologs from other organisms recognize different PAM sequences, expanding the targetable sequence space [31].
3.2 Mechanism of DNA Cleavage
The molecular mechanism of CRISPR-Cas9-mediated genome editing can be conceptually divided into three stages: target recognition, R-loop formation and conformational activation, and DNA cleavage [32]. In the first stage, the Cas9–sgRNA complex interrogates the genome by transiently binding to double-stranded DNA at PAM-containing sites. PAM recognition by the PI domain of Cas9 is the initiating event — it allows local DNA melting and facilitates directional scanning of the adjacent sequence by the sgRNA spacer [33].In the second stage, if the target DNA sequence is sufficiently complementary to the sgRNA spacer (typically requiring near-perfect complementarity over a 10–12 nucleotide "seed" region immediately adjacent to the PAM), stable hybridization occurs to form an R-loop, in which the sgRNA displaces the non-template DNA strand [34]. R-loop formation induces large conformational changes in Cas9 that reposition the HNH and RuvC domains into catalytically active configurations. In the third stage, the activated HNH domain cleaves the template strand, and the RuvC domain cleaves the non-template strand, generating a blunt-ended DSB 3 bp upstream of the PAM [29]. A key feature of this mechanism is that sgRNA–DNA mismatches at positions distal to the PAM (positions 1–8 of the spacer from the 5' end) are better tolerated than mismatches in the seed region, explaining the positional dependence of off-target effects [30].
3.3 DNA Repair Pathways and Editing Outcomes
Following Cas9-induced DSB formation, the cell employs one of two principal DNA repair pathways to resolve the break [35]. Non-homologous end joining (NHEJ) is the predominant and most rapid pathway in most cell types; it ligates the broken DNA ends without a homologous template, frequently introducing small insertions or deletions (indels) at the break site. When these indels occur within a coding exon, they commonly cause frameshifts resulting in premature stop codons and loss of gene function — a mechanism widely exploited for gene knockout studies [36]. Microhomology-mediated end joining (MMEJ), an error-prone alternative, produces predictable deletions templated by short flanking microhomologies and has been harnessed for precise indel engineering [37].Homology-directed repair (HDR), the second major pathway, requires a donor DNA template carrying sequences homologous to the sequences flanking the DSB. When a donor template is provided alongside the CRISPR-Cas9 components, HDR can mediate precise editing — including point mutations, small sequence insertions, or large fragment insertions — at the target locus [38]. However, HDR is active predominantly in dividing cells during the S and G2 phases of the cell cycle and is typically several-fold less efficient than NHEJ, limiting its practical utility in post-mitotic cells and tissues [39]. Strategies to enhance HDR efficiency, including cell cycle synchronisation, small molecule inhibitors of NHEJ, and optimised donor template designs, remain an active area of research [40].
3.4 Specificity, Off-Target Effects, and Engineering Improvements
A central concern in the application of CRISPR-Cas9 for therapeutic and research purposes is the potential for off-target cleavage at non-intended genomic loci. Off-target effects arise when the sgRNA hybridises to sequences with limited mismatches relative to the intended protospacer, leading to unintended DSBs that may cause mutations, chromosomal rearrangements, or, in therapeutic contexts, oncogenic transformation [41]. The frequency and pattern of off-target events are influenced by sgRNA design, Cas9 protein concentration, and the degree of chromatin accessibility at candidate off-target sites [42].Multiple strategies have been developed to detect and mitigate off-target effects. Unbiased genome-wide off-target detection methods — including GUIDE-seq, CIRCLE-seq, and DISCOVER-seq — allow comprehensive profiling of Cas9 cleavage sites across the genome [43]. To improve specificity, researchers have engineered high-fidelity Cas9 variants, including SpCas9-HF1 [44], eSpCas9 [45], and HypaCas9, which carry mutations that reduce non-specific DNA contacts while maintaining robust on-target activity. Paired nickase strategies, in which two Cas9 variants (each with one nuclease domain inactivated) introduce nicks on opposite DNA strands, require simultaneous engagement of both sgRNAs for DSB formation, substantially improving specificity at the cost of reduced flexibility [46]. Anti-CRISPR proteins, naturally occurring inhibitors of Cas systems, have additionally been explored as tools for temporal and spatial control of Cas9 activity [47].
4. Applications of CRISPR-Cas9 Technology
4.1 Applications in Agriculture
CRISPR-Cas9 technology holds transformative promise for agriculture by enabling rapid, precise, and targeted modification of crop and livestock genomes in ways that transcend the capabilities of conventional breeding or earlier transgenic approaches [14]. Unlike traditional genetic modification, which typically involves random integration of foreign DNA, CRISPR-Cas9 can be used to introduce targeted mutations — including gene knockouts, gene insertions, and precise base changes — without the incorporation of exogenous genetic material, a distinction that has significant implications for regulatory classification and public acceptance [48].Crop improvement represents the most intensively developed agricultural application. Researchers have employed CRISPR-Cas9 to enhance disease resistance, yield, nutritional quality, and abiotic stress tolerance in a wide range of economically important crops. In a seminal study, Shan et al. demonstrated targeted modification of the rice genome using CRISPR-Cas9 to generate plants with enhanced resistance to bacterial blight caused by Xanthomonas oryzae, by editing a susceptibility gene in the OsSWEET family [49]. Similar approaches have been applied to engineer resistance to powdery mildew in wheat [50], late blight in potato [51], and citrus canker in oranges [52]. The ability to simultaneously edit multiple genes — multiplexed editing — has further expanded the scope of agricultural applications, enabling simultaneous improvement of several traits within a single generation [53].
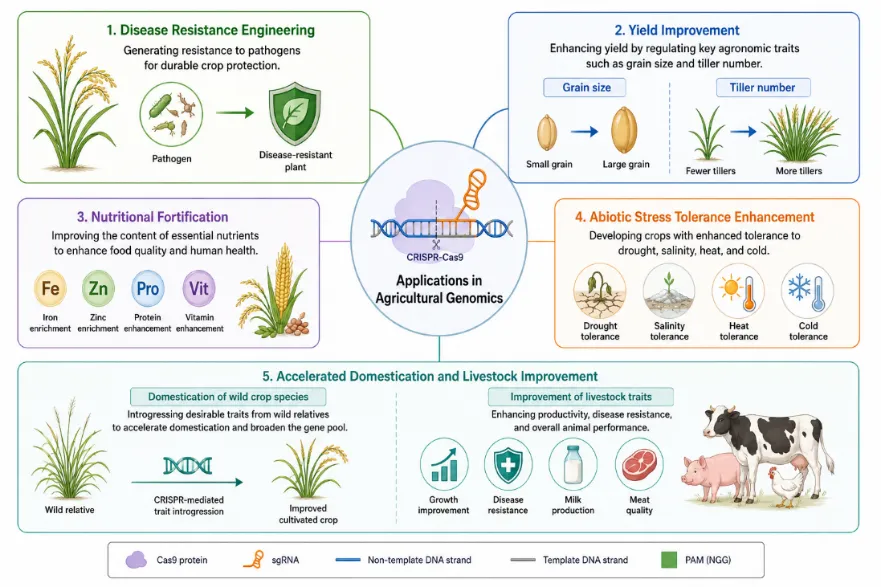
Figure 2: Applications of CRISPR-Cas9 technology in agricultural genomics
Yield improvement has been a particularly prominent target. Li et al. used CRISPR-Cas9 to target genes associated with grain size regulation in rice, achieving significant increases in yield without detrimental effects on agronomic performance [54]. More recently, CRISPR-Cas9 has been employed to modify multiple negative regulators of grain yield simultaneously, resulting in additive improvements across yield components [55]. In tomato, CRISPR-Cas9-mediated modification of genes controlling fruit size, plant architecture, and inflorescence branching has demonstrated the potential to rapidly domesticate wild tomato relatives with superior stress tolerance by recapitulating the genetic changes associated with historical crop domestication [56].
Nutritional enhancement represents another major avenue of agricultural CRISPR research. The technology has been applied to increase the content of health-promoting compounds — including anthocyanins, gamma-aminobutyric acid (GABA), lycopene, and vitamin content — and to reduce anti-nutritional factors such as acrylamide precursors and allergens in staple crops [57]. CRISPR-Cas9 has additionally been used to develop waxy corn and high-oleic soybean varieties with improved oil quality profiles that have advanced to regulatory approval and commercial development in the United States [58].In the livestock sector, CRISPR-Cas9 has been applied to edit genes associated with disease resistance, growth, reproduction, and product quality. Notable examples include the generation of pigs resistant to porcine reproductive and respiratory syndrome virus (PRRSV) by targeted disruption of the CD163 receptor gene [59], and the production of hornless cattle through editing of the polled locus, which could improve animal welfare without the welfare costs associated with traditional dehorning practices [60]. Aquaculture applications have also been explored, with CRISPR-Cas9 used to produce sterile salmon and triploid fish for containment of genetically modified fish, and to engineer disease-resistant shrimp strains [61].Despite this remarkable progress, the agricultural deployment of CRISPR-Cas9 faces substantial challenges. Regulatory frameworks governing CRISPR-edited crops vary substantially across jurisdictions: the United States has adopted a generally permissive approach for edits that could have arisen through conventional mutagenesis, while the European Union has until recently classified CRISPR-edited crops under its stringent GMO regulatory framework, a position being revised in ongoing legislative deliberations [62]. Public perceptions of CRISPR-edited food products — shaped by historical attitudes toward GMOs, media framing, and levels of scientific literacy — also influence acceptance and adoption [63].
4.2 Applications in Medicine
The medical applications of CRISPR-Cas9 have attracted extraordinary scientific and public attention since the technology's adaptation to mammalian cells. In the biomedical research context, CRISPR-Cas9 has fundamentally transformed the toolkit available for studying gene function, modelling disease, and discovering therapeutic targets [64]. By enabling rapid and scalable generation of gene knockouts — individually or in genome-wide pooled libraries — CRISPR-Cas9 has enabled functional genomic screens of unprecedented scope, identifying genes essential for cancer cell survival [65], drug resistance mechanisms [66], and host factors required for viral infection [67].The creation of disease models represents another transformative medical application. Prior to CRISPR-Cas9, the generation of genetically accurate disease models — particularly those carrying patient-specific mutations — was laborious, expensive, and limited to a small number of model organisms. CRISPR-Cas9 has enabled the rapid creation of patient-specific induced pluripotent stem cell (iPSC) disease models carrying disease-causing mutations, which can be differentiated into relevant cell types for mechanistic studies and drug screening [68]. For example, CRISPR-Cas9 was used to introduce disease-specific mutations into iPSCs to generate models of autosomal dominant polycystic kidney disease (ADPKD), enabling detailed study of the PKD2 gene's role in disease pathogenesis [69]. Similar approaches have been applied to model neurological diseases, cardiac conditions, and rare metabolic disorders [70].
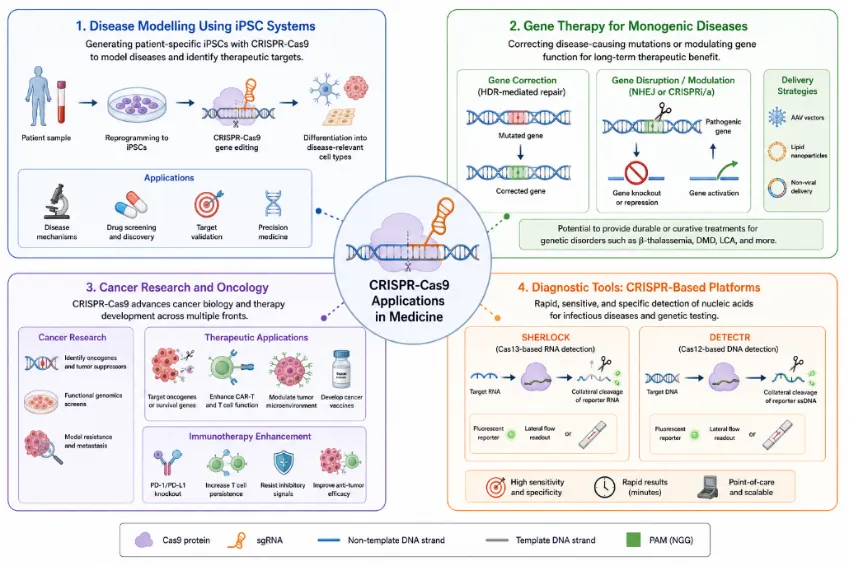
Figure 3: Overview of CRISPR-Cas9 applications in medicine. The diagram categorises current and emerging medical applications into four principal domains: (1) disease modelling using iPSC systems; (2) gene therapy approaches for monogenic diseases; (3) cancer research and oncology, including immunotherapy enhancement; and (4) diagnostic tools including SHERLOCK and DETECTR platforms.
The development of gene therapies — particularly for monogenic diseases caused by well-characterised mutations — represents perhaps the most clinically impactful application of CRISPR-Cas9. The technology offers the possibility of correcting disease-causing mutations directly in patient cells or tissues, potentially achieving durable therapeutic benefit from a single treatment. Early clinical-stage efforts have focused on blood disorders including sickle cell disease and beta-thalassemia, for which CRISPR-Cas9-mediated reactivation of fetal haemoglobin (HbF) has emerged as a particularly promising strategy [71]. In 2023, the FDA approved Casgevy (exa-cel), the first CRISPR-based therapeutic — a milestone marking the entry of CRISPR gene editing into mainstream clinical medicine [72].In the oncology domain, CRISPR-Cas9 has been applied to enhance the anti-tumour efficacy of T cell-based immunotherapies, including chimeric antigen receptor T (CAR-T) cell therapies. Multiplexed CRISPR editing has been employed to knock out immune checkpoint genes (such as PDCD1 encoding PD-1) and T cell receptor components in autologous and allogeneic T cells, with early- phase clinical trials demonstrating feasibility and acceptable safety [73]. CRISPR-Cas9 has also been used to identify novel cancer vulnerabilities through genome-wide synthetic lethality screens, revealing gene pairs whose simultaneous loss is selectively lethal in cancer cells, providing a rational basis for combination therapy strategies [74].CRISPR-based diagnostic platforms represent an innovative extension of the technology beyond its genome-editing function. Exploiting the collateral cleavage activity of Cas13 (and, to a lesser extent, the non-specific single-stranded DNA cleavage activity of activated Cas12a), researchers have developed rapid nucleic acid detection assays — SHERLOCK (Specific High-sensitivity Enzymatic Reporter unLOCKing) and DETECTR (DNA Endonuclease-Targeted CRISPR Trans Reporter) — capable of detecting viral infections, bacterial pathogens, and cancer-associated mutations from clinical samples with attomolar sensitivity [75]. These platforms were rapidly deployed during the COVID-19 pandemic to develop SARS-CoV-2 diagnostic tests with point-of-care potential [76].
4.3 Applications in Clinical Sciences
The translation of CRISPR-Cas9 technology from bench to bedside has proceeded with remarkable speed in the context of monogenic diseases and haematological conditions. Clinical research programmes have documented the safety and preliminary efficacy of CRISPR-Cas9-based interventions in human patients for the first time. A landmark early-phase clinical trial demonstrated that CRISPR-Cas9-edited T cells from patients with advanced, refractory cancers could be safely administered, with engineered cells persisting for over nine months without significant adverse effects [73,77].Cardiovascular disease represents another major focus of clinical CRISPR research. The PCSK9 gene, encoding a protease that degrades LDL receptors and thereby elevates plasma LDL cholesterol, has been identified as a compelling therapeutic target. In vivo delivery of CRISPR-Cas9 components targeting PCSK9 in the liver — using lipid nanoparticle delivery systems — produced sustained reductions in circulating LDL cholesterol in non-human primate models, with a single treatment achieving effects comparable to lifelong statin therapy [78]. Phase 1 clinical trials of in vivo CRISPR-based PCSK9 editing in patients with familial hypercholesterolaemia are ongoing, with early results indicating durable target engagement and a favourable safety profile [79].In infectious disease, CRISPR-Cas9 has been investigated as a strategy for targeting integrated viral genomes in latently infected cells — a major obstacle to HIV cure efforts. Disruption of the CCR5 co-receptor gene, which is required for HIV-1 entry into CD4+ T cells, has been explored as an ex vivo gene therapy approach to confer resistance to HIV infection in haematopoietic stem cells [80]. Separately, researchers have investigated CRISPR-Cas9-mediated excision of the latent HIV provirus from infected cells, with proof-of-concept studies in humanised mouse models demonstrating significant reductions in viral load [81].The principal outstanding challenges in clinical translation of CRISPR-Cas9 relate to the efficiency and safety of in vivo delivery — particularly to tissues and organs other than the liver — to the accurate characterisation and minimisation of off-target editing in patient cells, and to the long-term durability and reversibility of therapeutic effects. Substantial regulatory, manufacturing, and economic barriers also remain to be overcome to achieve broad patient access to CRISPR-based therapies [82].
4.4 Applications in Environmental Sciences
The application of CRISPR-Cas9 in environmental science has opened new possibilities for conservation biology, ecological management, and environmental monitoring [83]. While many of these applications remain at relatively early stages of development, they illustrate the broad potential reach of CRISPR-based tools beyond the laboratory and clinic.Gene drives — genetic systems that use CRISPR-Cas9 to spread engineered traits through wild populations at super-Mendelian frequencies — have attracted intense interest as tools for controlling mosquito-borne diseases. CRISPR-based gene drives targeting female fertility genes in Anopheles gambiae mosquitoes have demonstrated the ability to suppress laboratory cage populations to near extinction within a limited number of generations [84]. Analogous approaches are being developed for control of Aedes aegypti, the primary vector of dengue, Zika, and chikungunya viruses [85]. The ecological implications of releasing self-propagating gene drives into wild populations — including the potential for unintended spread to related species or for reversal of the intended trait — have prompted intense ethical and regulatory deliberation, with most researchers advocating for the development of geographically confined or reversible drive systems [86].Conservation biology represents another emerging area of CRISPR application. Researchers have proposed using CRISPR-Cas9 to introduce genetic variants associated with resilience — for example, to white-nose syndrome in North American bats or to thermal stress in coral reef organisms — into endangered populations to improve their survival prospects in the face of environmental change [87]. De-extinction efforts — aiming to resurrect characteristics of extinct species in closely related extant relatives — have also employed CRISPR-Cas9, most notably in the ongoing Colossal Biosciences project aimed at resurrecting traits of the woolly mammoth in the Asian elephant genome [88]. While these applications excite considerable scientific and public imagination, they raise profound questions about the ethics of intentional modification of wild populations, ecological risk, and the allocation of conservation resources [89].In the context of invasive species management, CRISPR-Cas9-based gene drives offer a theoretical mechanism to suppress or eliminate established invasive populations without sustained human intervention, potentially overcoming the limitations of conventional control methods. However, the irreversibility of gene drive deployment and the potential for cross-species gene flow have made regulatory and governance bodies highly cautious about authorising field trials [90]. Environmental monitoring applications of CRISPR diagnostics — using Cas12a or Cas13-based platforms to detect specific pathogens, pollutants, or indicator species in environmental samples — offer a more immediately deployable and lower-risk application of CRISPR technology in ecological management [91].
4.5 Applications in Nanotechnology
The intersection of CRISPR-Cas9 technology with nanotechnology has generated a synergistic research frontier with important implications for targeted delivery, imaging, and biosensing [92]. A major technical bottleneck in the therapeutic application of CRISPR-Cas9 is the challenge of efficiently and safely delivering the Cas9 protein and sgRNA into target cells and tissues in vivo. Nanotechnology-based delivery systems — including lipid nanoparticles (LNPs), polymeric nanoparticles, gold nanoparticles, and virus-like particles — have emerged as leading platforms for CRISPR component delivery, offering the potential to achieve tissue-specific targeting while reducing immunogenicity and systemic toxicity [93].Lipid nanoparticles, the most clinically advanced delivery platform for nucleic acid therapeutics (as validated by their use in COVID-19 mRNA vaccines), have been adapted to deliver Cas9 mRNA and sgRNA to the liver with high efficiency [94]. Engineering of LNP surface chemistry — through the incorporation of targeting ligands such as GalNAc or antibodies against cell-specific receptors — is enabling extrahepatic tissue targeting to organs including the lung, spleen, bone marrow, and tumours [95]. Gold nanoparticles conjugated to CRISPR components have demonstrated promise for local delivery to tumours, taking advantage of the enhanced permeability and retention (EPR) effect [96]. Exosome-based delivery vehicles — biologically derived nanoparticles with inherent cell-targeting capacity and low immunogenicity — have also been engineered to carry CRISPR-Cas9 components and are an area of active investigation [97].Beyond delivery, nanotechnology enables novel imaging and real-time tracking of CRISPR-Cas9 activity in cells and organisms. Nanoparticles functionalised with fluorescent dyes, quantum dots, or MRI contrast agents have been employed to monitor the biodistribution and intracellular trafficking of CRISPR components, providing insights into the kinetics of editing and facilitating optimisation of delivery protocols [98]. Nanotechnology-based biosensing platforms integrating CRISPR components — particularly the collateral cleavage activity of Cas12a and Cas13 — with gold nanoparticles, graphene electrodes, or plasmonic substrates have enabled ultrasensitive, label-free detection of nucleic acid targets for diagnostic applications [99]. These CRISPR-nanotech biosensors have demonstrated attomolarsensitivity with high specificity and are compatible with point-of-care deployment, suggesting broad utility in clinical diagnostics and environmental monitoring [100].
4.6 Applications in Bioinformatics and Biotechnology
The practical implementation of CRISPR-Cas9 experiments is intimately dependent on bioinformatics tools for sgRNA design, off-target prediction, and interpretation of editing outcomes from next-generation sequencing data [101]. A rapidly expanding ecosystem of computational tools has been developed to address these needs. sgRNA design platforms — including CRISPOR, Benchling, CHOPCHOP, and GuideScan — employ rule-based algorithms, machine learning models, and thermodynamic calculations to predict sgRNA on-target efficiency and off-target risk, facilitating the selection of optimal guide sequences for a given target locus [102]. Off-target prediction tools — including Cas-OFFinder, CRISPR-ML, and MOFF — model the permissive mismatches tolerated by Cas9 to generate ranked lists of potential off-target sites for experimental validation [103].The analysis of CRISPR editing outcomes from amplicon sequencing or whole-genome sequencing data requires dedicated computational pipelines. Tools such as CRISPResso2 and CRIS.py quantify the frequency and identity of indels and HDR events at target loci, enabling rigorous quality control and comparison of editing efficiencies across experimental conditions [104]. For genome-wide CRISPR screens — in which pooled sgRNA libraries are transduced into cells to simultaneously interrogate the functions of thousands of genes — dedicated analysis tools including MAGeCK, BAGEL2, and CRISPR-SURF are used to identify significant screen hits and delineate essential gene dependencies [105].In the biotechnology sector, CRISPR-Cas9 has been broadly adopted as a tool for metabolic engineering of microbial and mammalian cell lines for the production of biopharmaceuticals, industrial enzymes, biofuels, and specialty chemicals. The technology enables rapid, multiplexed genetic modification of production strains to optimise biosynthetic pathways, eliminate competing metabolic routes, and engineer cell lines with improved productivity, stability, and regulatory compliance [106]. CRISPR-Cas9 has also been applied to engineer cell lines used in the production of biologics — including CHO and HEK293 cells — to improve protein folding, glycosylation profiles, and secretion efficiency [107].
5. Discussion
5.1 Transformative Impact and Current Limitations
The foregoing review of CRISPR-Cas9 technology and its applications across multiple disciplines underscores the extraordinary impact of this platform on contemporary biology and medicine. Within barely a decade of its adaptation for eukaryotic genome editing, CRISPR-Cas9 has transformed experimental approaches across basic and applied life sciences, achieved first-in-class clinical approvals in gene therapy, and created entirely new research programmes in conservation biology, environmental management, and biosensing. It has democratised genome editing by radically lowering the technical and financial barriers to entry — enabling researchers in resource-limited settings and non-specialist laboratories to perform sophisticated genetic experiments — and has accelerated the pace of biological discovery across virtually every subdiscipline [108].Notwithstanding these achievements, several important technical limitations continue to constrain the full realisation of CRISPR-Cas9's potential. Off-target editing remains a concern in therapeutic contexts, where the consequences of unintended genomic modifications — particularly in long-lived post-mitotic cells or cells with oncogenic predisposition — require rigorous characterisation and monitoring [109]. The efficiency of HDR, essential for precise therapeutic correction of disease-causing mutations, remains suboptimal in most clinically relevant cell types. The size of the SpCas9 protein (approximately 4.2 kb coding sequence) approaches the packaging capacity of adeno-associated virus (AAV) vectors — the most widely used in vivo delivery vehicle — necessitating truncated forms, split intein strategies, or alternative smaller Cas proteins for AAV-based in vivo delivery [110].Immunogenicity represents an additional translational challenge. Pre-existing immune responses to Cas9 protein — which is derived from common commensal bacteria — have been detected in a substantial proportion of healthy human donors, raising concerns about immune clearance of edited cells or inflammatory responses to CRISPR components delivered in vivo [111]. Strategies to address this include engineering immunologically silent Cas9 variants, transient immunosuppression during delivery, and the use of Cas9 orthologues from organisms less commonly encountered by the human immune system [112].
5.2 Next-Generation CRISPR Tools
The limitations of first-generation CRISPR-Cas9 have spurred the development of a suite of next-generation CRISPR-based tools that expand the precision, versatility, and safety of genome editing. Base editors — including cytosine base editors (CBEs) and adenine base editors (ABEs) — use a catalytically impaired Cas9 (nickase or dead Cas9) fused to a deaminase enzyme to convert individual DNA bases (C to T, or A to G) within a defined editing window without inducing DSBs, substantially reducing the risk of undesired indels and large chromosomal rearrangements [113]. Base editors have been applied to correct pathogenic point mutations in cell lines, animal models, and, increasingly, in clinical development programmes.Prime editing, introduced by Anzalone et al. in 2019, further expands the scope of precise, DSB-free genome editing by enabling all twelve types of point mutation, as well as small insertions and deletions, to be installed at any genomic locus through the use of a reverse transcriptase fused to a Cas9 nickase and a prime editing guide RNA (pegRNA) that encodes both the targeting sequence and the desired edit [114]. Although prime editing efficiencies vary across cell types and genomic contexts, ongoing optimisation of the pegRNA scaffold, the reverse transcriptase domain, and the delivery format is rapidly improving its performance. Epigenome editing using catalytically dead Cas9 (dCas9) fused to chromatin-modifying enzymes — including DNA methyltransferases, histone methyltransferases, and histone deacetylases — enables durable, heritable modulation of gene expression without altering the underlying DNA sequence, offering a potentially reversible complement to classical genome editing [115].
5.3 Ethical Dimensions and Governance
The ethical controversies surrounding CRISPR-Cas9 are inseparable from its scientific development and must be addressed with rigour, inclusivity, and transparency. The most acute ethical concern — germline editing — pertains to the use of CRISPR-Cas9 to modify the DNA of human embryos, eggs, sperm, or early-stage embryos in ways that would be heritable by future generations. The birth in November 2018 of twin girls whose CCR5 gene had been edited as embryos by Chinese scientist He Jiankui — without adequate scientific justification, regulatory authorisation, or informed consent procedures — provoked near-universal condemnation from the global scientific and bioethics community and prompted renewed calls for an international moratorium on heritable human genome editing [116]. While most national regulatory bodies prohibit clinical heritable genome editing, the absence of a binding international treaty means that the risk of further illicit germline editing experiments cannot be ruled out [117].A fundamental question in the ethics of germline editing concerns the boundary between therapy and enhancement. While there is broad consensus that the correction of disease-causing mutations in embryos — were it proven sufficiently safe — might be ethically permissible in certain circumstances, the use of germline editing for non-therapeutic enhancement of traits such as intelligence, athleticism, or physical appearance raises profound concerns about social justice, discrimination, and the commodification of human genetics [118]. These concerns are amplified by the potential for germline editing to exacerbate existing inequalities if access to therapeutic genetic enhancement is restricted to affluent populations [119].Off-target effects in somatic gene therapy contexts — while less ethically fraught than germline editing — nonetheless require rigorous characterisation and transparent reporting to ensure patient safety and maintain public trust [120]. Intellectual property disputes over foundational CRISPR patents — most prominently the protracted litigation between the Broad Institute and the University of California — have raised legitimate questions about how patent ownership and licensing arrangements may influence the accessibility and affordability of CRISPR-based therapies and agricultural products, particularly in low- and middle-income countries [121].Responsible governance of CRISPR-Cas9 requires a multi-layered approach encompassing national regulatory frameworks, international coordination, professional self-regulation within the scientific community, and meaningful engagement with diverse public stakeholders. The Third International Summit on Human Genome Editing, held in 2023, reiterated that heritable human genome editing is not ready for clinical application and called for enhanced international oversight mechanisms [122]. The development of proportionate, adaptive governance frameworks capable of keeping pace with the rapid technical evolution of CRISPR tools represents one of the most pressing policy challenges of contemporary science [123].
6. CONCLUSION
CRISPR-Cas9 has established itself as one of the most consequential scientific innovations of the twenty-first century. Derived from an ancient bacterial immune mechanism, the system has been adapted into a remarkably versatile, precise, and accessible platform for genome editing that has fundamentally transformed biological research and created new therapeutic, agricultural, environmental, and industrial possibilities. The present review has synthesised the molecular basis of CRISPR-Cas9 function — from the structural features of the Cas9–sgRNA complex and the mechanism of target recognition and DNA cleavage, to the cellular repair pathways that determine editing outcomes — and has surveyed the expanding landscape of applications across agriculture, medicine, clinical sciences, environmental science, nanotechnology, bioinformatics, and biotechnology.In agriculture, CRISPR-Cas9 is accelerating the development of improved crop varieties and disease-resistant livestock, with significant implications for food security and agricultural sustainability. In medicine, it has progressed from a research tool for disease modelling and target discovery to an approved clinical therapeutic, with a growing pipeline of CRISPR-based gene therapies in clinical development. Environmental applications — spanning gene drives for disease vector control to conservation genomics and ecological monitoring — are advancing rapidly, though they require commensurate governance frameworks to manage their potentially profound ecological consequences. Synergies with nanotechnology are enabling more efficient and targeted delivery of CRISPR components, while CRISPR-based diagnostics are finding clinical and environmental applications enabled by the exquisite sensitivity of Cas effector collateral cleavage activities.Despite this extraordinary progress, significant technical and translational challenges remain, including the mitigation of off-target editing, the improvement of HDR efficiency, the development of effective in vivo delivery platforms for extrahepatic tissues, and the management of pre-existing and treatment-induced immune responses to Cas9. Next-generation tools — including base editors, prime editors, and epigenome editing platforms — are progressively addressing these limitations while expanding the precision and scope of programmable genome modification.The ethical, legal, and social dimensions of CRISPR-Cas9 technology demand sustained and inclusive engagement from scientists, clinicians, ethicists, policymakers, patient communities, and the broader public. The prohibition on heritable human genome editing must be robustly enforced through binding international mechanisms, while the development of somatic gene therapies must be pursued with rigorous safety evaluation, equitable access considerations, and transparent communication. The intellectual property landscape must evolve in ways that promote broad access to CRISPR-based innovations, particularly for applications of highest global health priority.In summary, CRISPR-Cas9 technology has ushered in a new era of precision biology with transformative implications across the life sciences and beyond. Its responsible development — guided by scientific rigour, ethical reflection, and inclusive governance — will be essential to realising the enormous promise of this technology while safeguarding against its potential harms. The continued integration of mechanistic insight, technological innovation, and thoughtful governance represents the most productive path toward harnessing CRISPR-Cas9 for the benefit of human health, food security, and environmental sustainability
REFERENCES:

Komal Gadage, Aniket Gadhave*, Shubham Kanawade, Clustered Regularly Interspaced Short Palindromic Repeats and Crispr-Associated Protein 9: Mechanisms, Applications, And Ethical Frontiers in Precision Genome Editing - A Comprehensive Review, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 7, 1909-1928. https://doi.org/10.5281/zenodo.21277967
 10.5281/zenodo.21277967
10.5281/zenodo.21277967