
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
1Research Scholar, Department of Pharmacology, Channabasweshwar Pharmacy College (Degree), Latur, Maharashtra, India 413512.
2Assistant professor, Department of pharmacology, Channabasweshwar Pharmacy College (Degree), Latur, Maharashtra, India 413512.
3Head of Department of Pharmacology, Channabasweshwar Pharmacy College (Degree), Latur, Maharashtra, India 413512.
4,5Research Scholar, Department of Pharmacology, Channabasweshwar Pharmacy College (Degree), Latur, Maharashtra, India 413512
Diabetes mellitus is a chronic metabolic disorder characterized by persistent hyperglycemia resulting from impaired insulin secretion, insulin resistance, or both. The pathophysiology of diabetes involves complex metabolic disturbances that contribute to the development of various complications affecting multiple organs. Increasing evidence suggests that oxidative stress plays a crucial role in the progression of diabetes by promoting the excessive generation of reactive oxygen species, which leads to cellular damage and metabolic dysfunction. In addition, chronic inflammation is considered an important factor in the development of insulin resistance through the activation of inflammatory signaling pathways and the release of pro-inflammatory cytokines. Due to the limitations and side effects associated with conventional antidiabetic drugs, medicinal plants have gained considerable attention as alternative therapeutic o its rich phytochemical composition, including lignans, flavonoids, and phenolic compounds that exhibit strong antioxidant activity. Although direct antidiabetic studies on this plant are limited, its antioxidant potential suggests that it may play a beneficial role in reducing oxidative stress associated with diabetes Therefore, further investigations are required to explore the pharmacological potential. and therapeutic applications of Magnolia liliflora in the management of diabetes.
Type 2 Diabetes Mellitus (T2DM) is a common metabolic disorder worldwide. It occurs mainly due to insufficient insulin secretion from pancreatic β-cells and decreased sensitivity of peripheral tissues to insulin. (1) Insulin secretion and its biological effects must accurately correspond to the body’s metabolic requirements. Therefore, the processes responsible for insulin production, its release, and its action in target tissues are strictly controlled. Any disruption in these regulatory mechanisms can result in metabolic disturbances, ultimately contributing to the development of Type 2 Diabetes Mellitus (T2DM).
1.1Pathophysiology of type 2 diabetes mellitus: -
In the pathophysiology of this condition, disruption of the regulatory feedback mechanisms linking insulin secretion and insulin action leads to elevated blood glucose levels.(2)
1.1.1 Insulin Resistance
Insulin resistance (IR) is defined as a reduced biological response of insulin-sensitive cells or tissues to circulating insulin, resulting in impaired glucose regulation (3) .It may arise due to three major mechanisms: decreased insulin secretion from pancreatic β-cells, the presence of circulating insulin antagonists (such as counter-regulatory hormones or other molecules that interfere with insulin signaling), and defective insulin action in peripheral target tissues (4). Insulin activity is modulated by several hormones. In the fed state, growth hormone and insulin-like growth factor-1 (IGF-1) influence insulin signaling, whereas during fasting, glucagon, glucocorticoids, and catecholamine’s counterbalance insulin effects to maintain glucose homeostasis. The insulin-to-glucagon ratio is crucial in regulating downstream metabolic pathways through enzyme phosphorylation. Excess levels of catecholamines and glucocorticoids enhance lipolysis, glycogenolysis, gluconeogenesis, and muscle protein breakdown, thereby contributing to the development of insulin resistance (5 ,6). Among extra-pancreatic tissues, skeletal muscle, adipose tissue, and the liver are the principal insulin-responsive organs. Impaired insulin action in these tissues often precedes systemic insulin resistance and ultimately contributes to the progression of Type 2 Diabetes Mellitus.
1.1.2. β-cell dysfunction
Both β-cell dysfunction and insulin resistance contribute to sustained hyperglycemia, a defining characteristic of type 2 diabetes mellitus. Genome-wide association studies (GWAS) have identified several susceptibility genes linked to type 2 diabetes that play important roles in regulating β-cell turnover and regeneration(7). Most genetic risk variants associated with type 2 diabetes in healthy individuals primarily affect insulin secretion by causing β-cell dysfunction rather than altering insulin action that leads to insulin resistance. This indicates that inherited defects in β-cell function, β-cell mass, or both play an important role in the early development of type 2 diabetes (8). Recent linkage analyses and genome-wide association studies (GWAS) have identified more than 40 genes that contribute to the risk of developing type 2 diabetes. Among these, the transcription factor 7-like 2 (TCF7L2) gene is considered the most significant susceptibility gene, increasing the risk of diabetes by approximately 1.7-fold(9). The potassium voltage-gated channel subfamily Q member 1 (KCNQ1) gene has been identified as a susceptibility gene for type 2 diabetes and is associated with impaired β-cell function and reduced insulin secretion. Several common genetic variants linked to neonatal diabetes mellitus and maturity-onset diabetes of the young (MODY), such as potassium inwardly rectifying channel subfamily J member 11 (KCNJ11), glucokinase (GCK), hepatocyte nuclear factor-4 alpha (HNF1α), and HNF1β, have also been identified as susceptibility variants associated with type 2 diabetes(10). Decreased expression of the transcription factor prospero homeobox 1 (PROX1), caused by cis-regulatory genetic variants, can impair insulin secretion from β-cells and thereby increase susceptibility to type 2 diabetes(11). β-cell dysfunction is considered a major factor in the development of type 2 diabetes and is often accompanied by insulin resistance (12). The relationship between β-cell dysfunction and insulin resistance is complex and closely interconnected. The development of hyperglycemia can further aggravate both β-cell impairment and insulin resistance. In many cases, β-cell dysfunction appears to play a more prominent role than insulin resistance in the progression of the disease.(figure 1)
|
Hyperglycaemia |
|
Beta cell dysfunction |
|
Insulin resistant |
|
Hyperglycaemia |
|
Type 2 diabetes |
FIGURE 1: Hyperglycemia-induced beta cell dysfunction, insulin resistance, and type 2 diabetes.
1.1.3. Hepatic Insulin Resistance:
Hepatic insulin resistance is a major factor contributing to hyperglycemia in Type 2 diabetes mellitus. It occurs when the liver fails to effectively suppress glucose production in response to insulin. Under normal physiological conditions, insulin reduces hepatic glucose production by inhibiting both gluconeogenesis and glycogenolysis, while simultaneously enhancing glucose uptake and its storage in the form of glycogen. (13)
Excessive hepatic gluconeogenesis is considered a major factor responsible for fasting hyperglycemia in patients with Type 2 diabetes mellitus. Evidence from studies using isotopic tracer techniques indicates that the increased hepatic glucose production observed in T2DM is mainly due to enhanced gluconeogenesis rather than glycogenolysis. In individuals with T2DM, the rate of gluconeogenesis may increase by approximately 30–50% compared with healthy subjects, contributing significantly to the elevated hepatic glucose output. (14). Various factors are responsible for the increased gluconeogenesis observed in hepatic insulin resistance. When the liver becomes resistant to insulin, its normal ability to suppress gluconeogenic pathways during the fed state is impaired, leading to their inappropriate activation. Consequently, major gluconeogenic enzymes such as phosphoenolpyruvate carboxykinase (PEPCK), glucose-6-phosphatase (G6Pase), and fructose-1,6-bisphosphatase remain abnormally active even in the presence of high insulin levels, resulting in increased hepatic glucose production in Type 2 diabetes mellitus (15). (Figure 2) The regulation of gluconeogenesis at the transcriptional level involves several important transcription factors and coactivators. Among them, peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PGC-1α) plays a central role by stimulating the expression of key gluconeogenic genes such as PEPCK and G6Pase. In conditions of hepatic insulin resistance, impaired insulin signaling leads to persistent activation of PGC-1α, which results in continuous stimulation of gluconeogenesis and contributes to increased glucose production in Type 2 diabetes mellitus (16). Forkhead box protein O1 (FoxO1) is another important transcription factor involved in the regulation of gluconeogenesis. Under normal physiological conditions, insulin activates Akt, which phosphorylates FoxO1 and causes its exclusion from the nucleus, thereby suppressing its activity. However, in hepatic insulin resistance, defective Akt signaling prevents this inactivation, allowing FoxO1 to remain active within the nucleus. As a result, it enhances the expression of gluconeogenic genes, contributing to increased glucose production in Type 2 diabetes mellitus(17).
Hepatic glycogenolysis plays an important role in maintaining glucose production, especially during fasting and in the early postprandial phase. In patients with Type 2 diabetes mellitus, impaired suppression of glycogenolysis along with defective regulation of glycogen synthesis contributes to hepatic insulin resistance (18). Glycogen phosphorylase (GP), the key rate-limiting enzyme responsible for glycogen breakdown, is normally inhibited by insulin through dephosphorylation. However, during hepatic insulin resistance, defective insulin signaling results in persistent activation of GP, causing continuous glycogen degradation even when glucose and insulin levels are elevated (18). In contrast, the activity of glycogen synthase (GS), the enzyme responsible for glycogen formation, is reduced due to impaired insulin signaling, which ultimately decreases the liver’s ability to store glycogen (18). Hepatic glycogen metabolism is regulated through complex interactions between hormonal signals and enzyme-mediated pathways. Glucagon, which is frequently elevated in type 2 diabetes, activates protein kinase A (PKA), promoting phosphorylation and activation of GP while simultaneously phosphorylating and inhibiting GS. This disruption in the normal balance between insulin and glucagon further aggravates glycogenolysis and contributes to excessive hepatic glucose production (19).
1.1.4 Mechanisms of lipid-induced hepatic insulin resistance:
Accumulation of intrahepatic lipids, particularly diacylglycerol (DAG), is considered a major factor linking non-alcoholic fatty liver disease to hepatic insulin resistance. DAG activates protein kinase C epsilon (PKCε), which interferes with insulin signaling by phosphorylating insulin receptor substrate-2 (IRS-2) in hepatocytes (20). Human studies have shown that the level of hepatic diacylglycerol (DAG) is a major determinant of insulin resistance, explaining nearly 64% of the variation observed in insulin sensitivity. The relationship between hepatic DAG content and insulin resistance (r = 0.80) is significantly stronger than the correlations reported with other lipid metabolites or inflammatory markers. Furthermore, DAG mainly accumulates within cytoplasmic lipid droplets in hepatocytes, and its increased concentration is closely associated with the activation of protein kinase C epsilon (PKCε) (20) (Figure 2). Another group of lipids, ceramides, also contributes to hepatic insulin resistance by activating protein phosphatase-2A (PP2A), which inhibits Akt signaling and disrupts insulin action. Ceramides are produced from saturated fatty acids such as palmitate, which are often elevated in obesity (21).
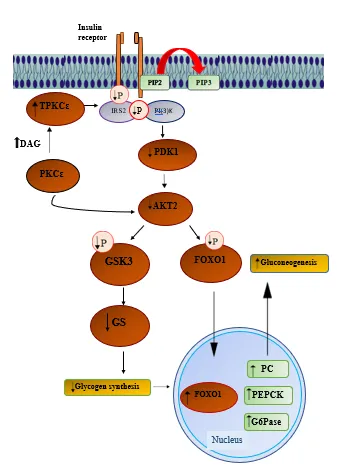
Figure 2: Molecular mechanism of diacylglycerol-PKCε-mediated hepatic insulin resistance
2.Role of oxidative stress:
Oxidative stress occurs when there is an imbalance between the generation of reactive oxygen species (ROS) and the capacity of the biological system to neutralize them or repair the damage caused by them. ROS are oxygen-containing free radicals, and the major biologically relevant species include superoxide anion (O₂⁻), hydrogen peroxide (H₂O₂), and hydroxyl radicals (OH•). To counteract these reactive molecules, the body possesses several antioxidant defense mechanisms. These include antioxidant vitamins such as vitamins A, C, and E; compounds like glutathione, α-lipoic acid, carotenoids, and coenzyme Q10; as well as antioxidant minerals including copper, zinc, manganese, and selenium. In addition, enzymatic antioxidants such as superoxide dismutase, catalase, glutathione peroxidase, and glutathione reductase play a crucial role in protecting cells from oxidative damage in conditions such as Type 2 diabetes mellitus . Antioxidants act together in a coordinated manner to neutralize different types of free radicals. When oxidative stress occurs, it can harm various cellular components, including proteins, membrane lipids, and nucleic acids (22). Oxidative stress is elevated in patients with diabetes, particularly in those with poor glycemic control (23). Studies have shown that exposure of aortic endothelial cells to high glucose levels increases reactive oxygen species (ROS) production by approximately 250% within 24 hours, which subsequently leads to a 330% increase in lipid peroxidation within 168 hours (24). Experimental studies in animals have demonstrated that inhibition of oxidative stress using certain antioxidants can help prevent the development of diabetic complications (25–27). However, clinical studies in humans using antioxidants such as α-tocopherol and ascorbic acid have shown limited benefits (28,29). In contrast, supplementation with α-lipoic acid has been reported to improve neuropathic symptoms in patients with diabetic distal symmetric polyneuropathy (30–32). Therefore, developing more targeted antioxidant therapies based on the mechanisms of diabetes-induced oxidative stress may be beneficial for preventing diabetic complications.
2.1 The Mechanisms Of Oxidative Stress In Diabetes:-
Several mechanisms have been proposed to explain diabetes-induced oxidative stress (Figure 3). One important source is glucose auto-oxidation, in which glucose in its enediol form undergoes transition-metal–dependent oxidation, producing superoxide radicals that can subsequently form hydrogen peroxide and highly reactive hydroxyl radicals if not detoxified by antioxidant enzymes such as catalase or glutathione peroxidase (33). Another mechanism involves activation of the polyol pathway, where the enzyme aldose reductase converts excess intracellular glucose into sorbitol using NADPH as a cofactor. During hyperglycemia, increased consumption of NADPH may reduce the availability of reduced glutathione, thereby weakening cellular antioxidant defense systems (34). In addition, hyperglycemia activates protein kinase C (PKC) through increased synthesis of diacylglycerol, which stimulates ROS generation via activation of NAD(P)H oxidase in vascular cells (35). NAD(P)H oxidase is present in various cell types, including endothelial and smooth muscle cells, where it contributes to superoxide production (36). Furthermore, hyperglycemia enhances non-enzymatic glycation reactions, leading to the formation of advanced glycation end products (AGEs), which promote oxidative stress through activation of AGE receptors and increased ROS production (37). Glycation can also impair antioxidant enzymes such as superoxide dismutase, further aggravating oxidative damage (38).
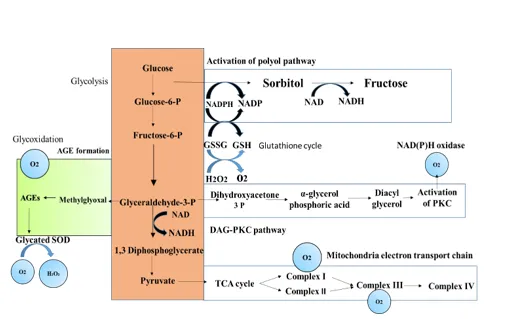
Figure 3 :- Proposed mechanism leading to diabetes-induced oxidative stress in cells
Role of Inflammation: -
Oxidative stress (OS) stimulates the production of inflammatory mediators, which further promote the generation of reactive oxygen species (ROS). Excess ROS can harm cells by damaging essential cellular components such as proteins, lipids, and DNA (39). Persistent hyperglycemia is known to enhance free radical formation and weaken endogenous antioxidant defense mechanisms through multiple pathways (40). ROS include a wide range of oxidant molecules that participate in cellular signaling as well as oxidative injury. These oxidants are generally classified into radical (one-electron) and non-radical (two-electron) forms. Radical ROS include superoxide anion (O₂•⁻), hydroxyl radical (HO•), peroxyl radical (ROO•), and alkoxyl radical (RO•), which are capable of inducing significant biomolecular damage during oxidative reactions. Non-radical ROS include molecules such as hydrogen peroxide (H₂O₂), singlet oxygen (¹O₂), triplet carbonyls, and ozone (O₃), each contributing differently to cellular processes ranging from signaling to oxidative injury (41). Moreover, ROS are closely involved in the regulation of inflammation and metabolic pathways. Excessive ROS generation can impair insulin action and insulin secretion, thereby promoting insulin resistance and creating a vicious cycle of metabolic dysfunction (42). Figure . This review will focus on the relationship between inflammation and IR, and we analyze the mechanisms relating to how inflammatory cytokines, signaling pathways.
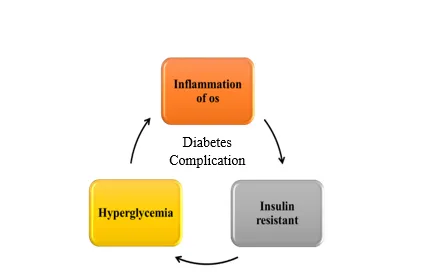
Figure 4. The vicious cycle of inflammation and hyperglycaemia
3.1Cytokines That Link Inflammation to IR:-
3.1.1. TNF-𝛼. :- Early studies conducted in the 1990s identified a strong association between inflammation and insulin resistance (IR), particularly focusing on the role of tumor necrosis factor-α (TNF-α) [43]. TNF-α is a pro-inflammatory cytokine mainly released from adipose tissue and contributes to insulin resistance by stimulating lipolysis in adipocytes and increasing the serine/threonine phosphorylation of insulin receptor substrate-1 (IRS-1) [44,45]. Several signaling pathways are involved in this process, including the IKKβ/NF-κB pathway, which plays an important role in the development of IR [46,47] (see Figure 5). TNF-α can influence glucose metabolism differently depending on adipose tissue type; it enhances glucose uptake in both visceral and subcutaneous adipocytes through activation of the AMP-activated protein kinase (AMPK) pathway, but it can also promote insulin resistance in visceral adipocytes via activation of the JNK1/2 pathway [48]. Due to these tissue-specific effects, therapeutic strategies targeting TNF-α signaling, such as the use of soluble tumor necrosis factor-like weak inducer of apoptosis (sTWEAK), have been explored to improve insulin resistance [49]. Furthermore, circulating levels of TNF-α are reported to be higher in males than in females and are elevated in obese individuals compared with lean individuals, suggesting a greater risk of insulin resistance and related disorders, including cardiovascular disease, particularly in obese men [50].
3.1.2. IL-1𝛽 :-Interleukin-1β (IL-1β) is an important pro-inflammatory cytokine whose secretion is regulated by inflammasome activation. It contributes to insulin resistance (IR) by disrupting insulin signaling in peripheral tissues and macrophages, which may reduce β-cell insulin sensitivity and impair insulin secretion [51,52]. During hyperglycemic conditions, the production of IL-1β increases in several cell types, including endothelial cells and monocytes [53]. IL-1β also plays a key role in initiating and sustaining inflammation-related organ dysfunction associated with type 2 diabetes mellitus (T2DM) [54] (see Figure 5). Furthermore, elevated IL-1β levels can promote systemic inflammation and inhibit insulin action in major insulin-target cells such as macrophages, thereby contributing to the progression of insulin resistance [55].
3.1.3. IL-6.:- Interleukin-6 (IL-6) is an inflammatory cytokine secreted by several tissues, especially adipose tissue, and it plays an important role in the development of insulin resistance (IR). IL-6 contributes to IR by decreasing the expression of glucose transporter-4 (GLUT-4) and insulin receptor substrate-1 (IRS-1), which are essential for proper insulin signaling. These effects occur mainly through activation of the Janus kinase–signal transducer and activator of transcription (JAK-STAT) signaling pathway and by increasing the expression of suppressor of cytokine signaling-3 (SOCS3) [56,57](see Figure 5). In addition, IL-6 can induce insulin resistance by inhibiting the phosphoinositide 3-kinase (PI3K) pathway and impairing glycogen synthesis through down-regulation of microRNA-200s (miR-200s) and up-regulation of GATA-2 (FOG-2) [58,59]. Furthermore, IL-6 stimulation has been associated with insulin resistance in human skeletal muscle by promoting toll-like receptor-4 (TLR-4) gene expression through STAT3 activation [60].
3.1.4. Leptin.:- Leptin is a hormone mainly produced by white adipose tissue (WAT) and plays an important role in regulating appetite and energy balance [61]. It reduces food intake and increases energy expenditure through neuronal signaling pathways. Leptin levels are influenced by nutritional status and are associated with the development of insulin resistance (IR) [62,63]. In obese individuals, leptin resistance is commonly observed, where high leptin levels fail to produce normal metabolic effects. Leptin signaling through the PI3K pathway is also important for glucose metabolism and pancreatic β-cell function [55–65]. Due to its effects on glucose, insulin sensitivity, and lipid metabolism, leptin is considered a potential biomarker and therapeutic target for insulin resistance [66–69].
3.1.5. Adiponectin: -Adiponectin is a hormone mainly produced by white adipose tissue (WAT), and its levels are usually reduced in conditions such as obesity, insulin resistance (IR), and type 2 diabetes mellitus (T2DM) [62,70]. It generally acts as an anti-inflammatory cytokine and plays an important role in regulating glucose metabolism. Adiponectin improves insulin sensitivity through two main receptors, AdipoR1 and AdipoR2. Activation of AdipoR1 stimulates the AMPK pathway, which suppresses hepatic gluconeogenesis and lipogenesis, whereas AdipoR2 activates PPAR-α signaling to enhance glucose utilization [71,72]. These receptors are mainly expressed in skeletal muscle and the liver, respectively, and together contribute to improving hepatic insulin resistance and metabolic regulation [52,73].
3.1.6. Resistin:- Resistin is an inflammatory hormone that is produced by adipocytes in rodents and mainly by macrophages in humans. Its levels increase along with inflammatory mediators and are associated with obesity and insulin resistance (IR) [74,75]. Resistin contributes to IR by promoting the expression of pro-inflammatory cytokines such as TNF-α and IL-6 through the NF-κB signaling pathway. It can also activate TLR4 receptors, which stimulate JNK and MAPK signaling pathways, thereby enhancing inflammation and insulin resistance [76].
3.1.7.MCP-1 :
Monocyte chemoattractant protein-1 (MCP-1) is a pro-inflammatory chemokine produced by adipocytes, macrophages, and endothelial cells. Its expression increases during obesity, especially in visceral adipose tissue, leading to the recruitment of macrophages and other immune cells that enhance inflammatory cytokine production [44,77,78]. MCP-1 contributes to insulin resistance by regulating inflammatory responses, lipid metabolism, and insulin sensitivity through signaling pathways such as ERK1/2 and p38 MAPK [79]. Its receptor, CCR2, also plays an important role in inflammation and metabolic dysfunction, indicating that MCP-1 is a key factor in the development of inflammation-induced insulin resistance [80].
Figure 5: Influence of the inflammatory cytokines on the status of insulin resistance
4. Signaling Pathways Linking Inflammation to Insulin Resistance: -
4.1.IKKβ/NF-κB Pathway: -The NF-κB pathway is an important regulator of inflammation and immune responses. Under normal conditions, NF-κB remains inactive in the cytoplasm by binding with IκB proteins. However, in response to metabolic stress or inflammatory stimuli such as obesity, the IKK complex becomes activated, leading to the phosphorylation and degradation of IκB. This process allows NF-κB to move into the nucleus and promote the expression of pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6 [43,81] (see figure 2 )Activation of the IKKβ/NF-κB pathway increases inflammation and contributes to insulin resistance, whereas inhibition of this pathway improves insulin sensitivity and glucose tolerance [83–85].
4.2.JNK Pathway:
The c-Jun N-terminal kinase (JNK) pathway is a member of the MAPK family and includes three isoforms: JNK-1, JNK-2, and JNK-3. This pathway plays an important role in inflammation, obesity, metabolic syndrome, and insulin resistance (IR) by regulating the production of pro-inflammatory cytokines and cellular stress responses [86–88]. JNK can also be activated by endoplasmic reticulum (ER) stress, which leads to serine phosphorylation of insulin receptor substrate-1 (IRS-1) (see figure 2 ) and disruption of insulin signaling. Increased JNK activity under diabetic conditions contributes to insulin resistance, whereas inhibition of JNK improves glucose tolerance and insulin sensitivity [89–91]. Additionally, excessive activation of JNK in insulin-sensitive tissues and pancreatic β-cells further promotes inflammation and insulin resistance [92–95].
|
Glycogen |
|
FOG-2 |
|
miR-200 |
The inflammasome is a cytosolic protein complex that regulates inflammatory responses by controlling the secretion of cytokines such as IL-1β and IL-18. It plays an important role in metabolic disorders, including obesity and insulin resistance [54,96]. One of the key components, NLRP3 inflammasome, is activated by metabolic stress and mitochondrial dysfunction, linking saturated fatty acids to chronic inflammation. Activation of inflammasomes can impair insulin sensitivity and promote macrophage infiltration into adipose tissue through the action of caspase-1 [96–99] (see figure 2 ). Studies have shown that reducing inflammasome activity improves insulin signaling, decreases inflammation, and enhances insulin sensitivity, suggesting that it may be an important therapeutic .
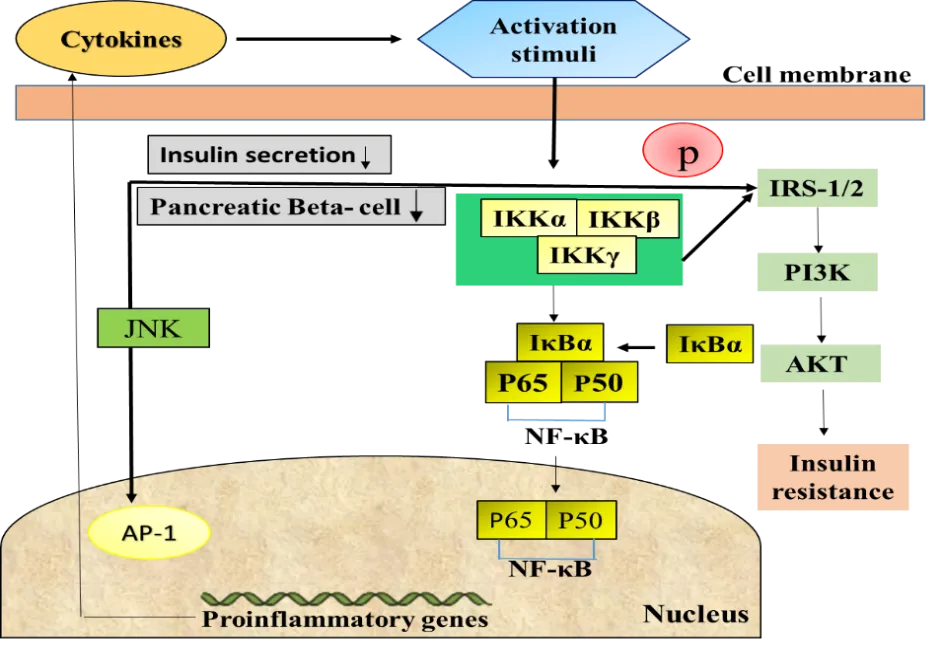
Figure 6: Inflammatory pathways linking inflammation to insulin
5.Role of Medicinal Plants in Diabetes Management: -
Medicinal plants have been widely utilized in traditional systems of medicine for the prevention and management of diabetes mellitus and its associated complications . A large number of plant species contain biologically active phytochemicals such as flavonoids, alkaloids, phenolic compounds, tannins, and terpenoids that exhibit significant antihyperglycemic effects [101,102,103]. These bioactive constituents regulate glucose metabolism through multiple mechanisms, including stimulation of insulin secretion from pancreatic β-cells, enhancement of insulin sensitivity in peripheral tissues, and increased glucose uptake in skeletal muscle and adipose tissue [101,103]. In addition, many medicinal plants possess strong antioxidant and anti-inflammatory activities, which play an important role in reducing oxidative stress and chronic inflammation that contribute to the development of insulin resistance and diabetes-related complications [102,104]. Several well-known medicinal plants such as Momordica charantia, Gymnema sylvestre, and Trigonella foenum-graecum have demonstrated promising antidiabetic potential in various experimental and clinical studies by improving glucose homeostasis and protecting pancreatic β-cells [103,105]. These plants may also inhibit carbohydrate-digesting enzymes, reduce hepatic glucose production, and modulate insulin signaling pathways, thereby contributing to better glycemic control [102,104]. Among the various medicinal plants investigated for their therapeutic potential, species belonging to the genus Magnolia lilliflora have recently attracted considerable scientific interest due to their rich phytochemical profile and diverse pharmacological activities, including antioxidant, anti-inflammatory, and possible antidiabetic effects [104,105].
6.Antioxidant Activity of Medicinal Plants:-
Oxidative stress plays a crucial role in the development and progression of diabetes mellitus and its related complications. It occurs when the production of reactive oxygen species (ROS) exceeds the capacity of the endogenous antioxidant defense system, leading to cellular damage and metabolic dysfunction [107,108]. Medicinal plants are considered an important source of natural antioxidants because they contain various bioactive compounds such as flavonoids, phenolic acids, tannins, and vitamins that can neutralize free radicals and reduce oxidative damage. These phytochemicals help in protecting pancreatic β-cells, improving insulin secretion, and enhancing glucose metabolism in diabetic conditions [100,101]. In addition, plant-derived antioxidants may inhibit lipid peroxidation, reduce oxidative stress-induced inflammation, and improve insulin sensitivity in peripheral tissues [108,101]. Therefore, medicinal plants with strong antioxidant properties are considered promising therapeutic agents for the prevention and management of diabetes and its complications [102].
7.Anti-inflammatory Activity of Medicinal Plants:-
Chronic inflammation plays a significant role in the development and progression of diabetes mellitus and insulin resistance [110,112]. In diabetic conditions, increased production of pro-inflammatory cytokines such as TNF-α, IL-6, and IL-1β activates inflammatory signaling pathways including NF-κB and JNK, which interfere with insulin signaling and glucose metabolism [112,113]. Persistent activation of these inflammatory mediators contributes to β-cell dysfunction, reduced insulin sensitivity, and metabolic complications associated with diabetes [113]. Medicinal plants contain numerous bioactive compounds such as flavonoids, phenolic acids, and terpenoids that exhibit strong anti-inflammatory properties [108]. These phytochemicals can suppress the production of inflammatory cytokines, inhibit NF-κB activation, and reduce inflammatory responses in metabolic tissues [108,107]. By modulating these inflammatory pathways, plant-derived compounds may improve insulin sensitivity and help prevent the progression of diabetes and its complications [,109]. Therefore, medicinal plants with potent anti-inflammatory activity are considered promising therapeutic agents in the management of diabetes.
Since oxidative stress and inflammation play a major role in the development and progression of diabetes, medicinal plants possessing antioxidant and anti-inflammatory properties have attracted significant attention for diabetes management. Numerous plant species have been reported to improve glucose metabolism and reduce diabetes-related complications through various bioactive phytochemicals. Among these medicinal plants, species belonging to the genus Magnolia have gained increasing scientific interest due to their diverse pharmacological activities. These plants are known to contain several biologically active compounds with antioxidant, anti-inflammatory, and potential antidiabetic effects. Therefore, the following section focuses on the photochemistry and antidiabetic potential of Magnolia.
8.Pharmacological Potential of Magnolia:-
Plants belonging to the genus Magnolia have been widely investigated for their diverse pharmacological properties, including antioxidant, anti-inflammatory, antimicrobial, and neuroprotective activities [114,115]. These biological effects are mainly attributed to the presence of various bioactive compounds such as lignans, flavonoids, and phenolic constituents [115]. Among the different species, Magnolia liliflora has attracted scientific interest due to its traditional medicinal uses and phytochemical richness [116]. Although limited studies have directly evaluated the antidiabetic potential of Magnolia liliiflora, several reports indicate that species of Magnolia exhibit strong antioxidant and anti-inflammatory activities, which are important in reducing oxidative stress and inflammation associated with metabolic disorders such as diabetes [,114,115].
9.Phytochemicals of Magnolia liliiflora:-
The medicinal properties of Magnolia liliiflora are mainly attributed to its diverse phytochemical constituents. Studies have reported that this plant contains several bioactive compounds including lignans such as magnolol and honokiol, along with flavonoids and various phenolic compounds [113]. These phytochemicals are known for their strong antioxidant and anti-inflammatory activities. Lignans such as magnolol and honokiol are considered the major active components of many species of the genus Magnolia and have been reported to exhibit multiple pharmacological effects including antioxidant, anti-inflammatory, and protective activities against oxidative stress [114]. In addition, flavonoids and phenolic compounds present in the plant contribute significantly to free radical scavenging activity and help reduce oxidative damage in biological systems [115]. Therefore, the presence of these phytochemicals suggests that Magnolia liliiflora may possess important therapeutic potential for the management of oxidative stress-related disorders.
10.Antioxidant Activity of Magnolia liliiflora: -
Oxidative stress plays a crucial role in the development of metabolic disorders including diabetes mellitus and its associated complications. Excessive generation of reactive oxygen species (ROS) can damage cellular components such as lipids, proteins, and DNA, leading to impaired metabolic functions . Plants belonging to the genus Magnolia are known to possess strong antioxidant activity due to the presence of bioactive compounds such as lignans, flavonoids, and phenolic constituents [113]. Among these compounds, magnolol and honokiol have been widely reported as potent natural antioxidants capable of scavenging free radicals and reducing oxidative stress in biological systems. Although specific antidiabetic studies on Magnolia liliiflora are limited, its phytochemical composition and antioxidant potential suggest that this plant may help in reducing oxidative damage associated with metabolic disorders [114]. Therefore, further experimental investigations are required to evaluate the antioxidant and possible antidiabetic potential of Magnolia liliiflora.
11.Conclusion and Future Perspectives: -
Diabetes mellitus is a complex metabolic disorder associated with oxidative stress and chronic inflammation, which contribute to the development of insulin resistance and related complications. Medicinal plants have attracted significant attention as potential therapeutic agents due to their diverse bioactive phytochemicals and pharmacological activities. Plants belonging to the genus Magnolia are known to contain several bioactive compounds such as magnolol, honokiol, flavonoids, and phenolic constituents that exhibit antioxidant and anti-inflammatory properties. Although limited studies have specifically investigated the antidiabetic potential of Magnolia liliiflora, its phytochemical composition and reported antioxidant activity suggest that this plant may have promising therapeutic potential in managing oxidative stress-related disorders. Therefore, further experimental and clinical studies are required to explore the antidiabetic mechanisms and pharmacological efficacy of Magnolia liliflora.
REFERENCES

Srushti Umbare, Balaji Ajgunde, Padmaja Giram, Madhuri Shinde, Priyanka Wadkar, Review on Diabetes Mellitus: Pathogenesis, Oxidative Stress, Inflammation and Herbal Bioactive Compounds, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 6, 895-915. https://doi.org/ 10.5281/zenodo.20530243
 10.5281/zenodo.20530243
10.5281/zenodo.20530243