
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
Mula Education Society's College of Pharmacy, Sonai, Newasa, Ahilyanagar 414105
High recurrence rates and a suboptimal pharmacological armamentarium are hallmarks of calcium oxalate nephrolithiasis, and current drugs (thiazide diuretics, potassium citrate and allopurinol) target individual factors of the metabolic matrix without affecting the physicochemical processes of nucleation, crystal growth, aggregation and renal tubular adhesion, which are responsible for stone formation. Bioactive compounds, however, obtained from plants have a structurally diverse and multi-targeted pharmacological action that can act at any step of this cascade. The present review encompasses the current knowledge of polyphenols, flavonoids, tannins, saponins, alkaloids and terpenoids, their calcium chelating, crystal surface adsorbing and aggregation inhibiting properties and in turn, the basis of their structures. As the cellular basis to looks at the molecular biology of crystal-induced renal injury, including the generation of reactive oxygen species by NADPH oxidases, activation of the NF-?B and NLRP3 inflammasome pathways and the cytoprotective Nrf2/antioxidant response element pathway. Data from in vitro crystallization assays, preclinical findings with ethylene glycol models, and emerging clinical data are critically evaluated against the pharmacokinetic constraints, such as poor oral bioavailability, high degree of hepatic conjugation, and concentration achievability gaps, that hamper direct translation of experimental findings. The most scientifically promising routes to further bridging this translational gap and progressing plant-derived crystallization modulators towards clinical proof-of-concept in nephrolithiasis prevention are identified as being nano-enabled delivery strategies and network pharmacology-based compound prioritisation.
Kidney stone disease is a uniquely challenging condition among the urological diseases; it is painful, expensive and often recurrent; and yet there is no single effective pharmaceutical treatment [1]. In humans, calcium oxalate is the most common lithogenic mineral (75%). The prevalence of nephrolithiasis in populations of European ancestry is estimated to be from 5 to 10% of adults and occurs approximately twice as often in men as in women under normal epidemiological conditions [2]. The high burden of the disorder on working population, which is compounded by climate-induced dehydration, obesity and metabolic syndrome, dietary transitions, has been proven in the GBD 2021 data; since 1990, the number of patients, cases and DALYs associated with urinary stones in the age group 20-54 has risen significantly across the globe [3]. It is not a single attack that makes this epidemiological course so difficult to treat, but the fact that the attacks tend to recur so much. The incidence of stone recurrence in established stone formers is 15%-20% per year with 27% to 50% cumulative at 5 years if no specific pharmacological treatment is applied [4].
There is a standard pharmacological armamentarium for preventing recurrent calcium oxalate stone formation: all of these drugs target one metabolic aspect of the risk of crystallization. Thiazide diuretics facilitate reabsorption of calcium in the tubules and have been found to decrease the recurrence of stones in some clinical trials, although a landmark NEJM trial showed that the dose required to achieve this effect should be carefully titrated, and a systematic review of the literature found evidence was directionally positive, albeit low-strength [5]. An acknowledged risk of allopurinol is an increased risk of acute kidney injury when compared with other drug classes (3.25-fold, according to analysis of the WHO adverse drug reaction database) and an increased risk of stone recurrence when compared with thiazides (32%, according to medical claims data). Thiazides and citrate must be co-administered with frequent potassium monitoring; liver enzyme monitoring is needed with allopurinol, which can present a monitoring burden that makes the drug difficult to adhere to in patients who are not particularly ill [6]. In addition to the safety issues, none of these agents acts at the basic level of the crystallization cascade the physical processes of nucleation, crystal growth and aggregation which determine whether a supersaturated urine will form a clinically relevant stone [7].
This therapeutic unmet need has maintained a continued scientific interest in the plant-derived bioactive compounds as multi-targeted alternative to the conventional prophylaxis. Medicinal plants have been used in traditional medicine system in South and Southeast Asian, Middle East and Latin American countries for centuries for the treatment of urinary stones and constitute an ethnopharmacological basis for the modern scientific study of antiurolithiatic activity; in many ways, such traditional use preceded and inspired the present-day research [8]. The pharmacological relevance of these plants is not only based on the empirical tradition, but also, on the structure and physicochemical properties of the phytochemicals, namely polyphenols, flavonoids, tannins, alkaloids and terpenoids, which have been reported in increasingly large experimental literature to directly interfere with calcium oxalate nucleation kinetics, modify crystal morphology, inhibit crystal aggregation, reduce the urinary supersaturation of calcium oxalate, and protect renal tubular epithelial cells from oxidative and inflammatory injury caused by crystal deposition. These compounds also seem to have convergent mechanisms of action affecting multiple steps of the stone formation cascade, as compared to conventional drugs that affect only a single metabolic parameter [9,10].
The present review summarizes the existing knowledge of the bioactive compounds of plants and their mode of action in the modulation of calcium oxalate crystallization. It consists of a sequence of lectures that gradually build the readers' knowledge from pathophysiology and crystallisation mechanics to the molecular biology of crystal-induced renal injury, the evidence base from in vivo and in vitro studies, and the importance of bioavailability issues for the credible translation of observed effects into real-world impact the "structural pharmacognosy" of antiurolithiatic compound activity. The aim of this book is to give pharmaceutical scientists and researchers in the area of natural product pharmacology a scientifically sound and mechanistic explanation of the present scenario and the most promising directions of research.
2. Pathophysiology of Calcium Oxalate Stone Formation: A Crystallization Perspective
The process of stone formation in the urinary tract is not a single event but is the result of a series of physicochemical and biological events, which take place in a given sequence in the nephron [11]. The main condition for urinary supersaturation is the condition of urinary calcium and oxalate ionic product exceeding the calcium oxalate solubility product under the urinary conditions of pH, ionic strength, and temperature [12]. Ironically, the condition of calcium oxalate being supersaturated in normal human urine is the rule, not the exception, but stones are the exception and don't form in the population in general. In physiological conditions, there is a complex endogenous system of crystallization inhibitors, which effectively inhibits the process of nucleation and subsequent crystal formation, thus resolving this apparent contradiction [13].
The initial event in stone formation is nucleation, which is the buildup of the smallest, thermodynamically stable cluster of mineral constituting the stone, either in the free solution or more likely, in the environment of the renal tubular lumen, on pre-existing surfaces like cellular debris, proteins, or already deposited mineral deposits [14]. When the initial nucleus is reached, crystal growth is achieved by the orderly deposition of calcium and oxalate ions onto the faces of the existing crystals, forming the characteristic shape of calcium oxalate monohydrate (COM) or the less clinically important calcium oxalate dihydrate (COD). Relative to the composition of the kidney stones, the ratio of COD to COM is found to be greater than 2:1 in the stone analysis and COM is the thermodynamically stable polymorph and accounts for the majority of mineral components of kidney stones in clinical practice [15]. This polymorphic distinction has also received considerable pathological significance: adhesive forces of COM crystals to the surface of renal epithelial cells have been shown to be significantly higher than those of COD crystals by atomic force microscopy, which can provide a molecular explanation for why COD-forming urine has a lower stone risk than COM-dominated crystalluria [16]. The process of crystal growth is followed by the physical aggregation (clustering) of individual microcrystals that leads to the formation of larger conglomerates, and it is the aggregation, not the crystal growth of the individual crystals that most directly determines whether a particle becomes large enough to obstruct the tubular lumen and begin the process of stone retention [17].
The interplay of crystallization promotion and inhibition determines whether the supersaturation is a clinical stone. Citrate is the most important inorganic inhibitor. Citrate interferes with calcium oxalate crystallization in two ways: citrate directly adsorbs to COM crystal step surfaces via its carboxylate functional groups, sterically preventing further ion deposition and citrate forms soluble complexes with calcium ions, which lowers the concentration of free calcium ions and thus the effective supersaturation. One of the most common and treatable metabolic abnormalities in nephrolithiasis is hypocitraturia (urinary citrate levels below 320 mg per day) [18]. Hypocitraturia has been seen in 20 to 60% of calcium stone formers. Magnesium also decreases urinary oxalate availability by forming complexes, and pyrophosphate decreases the growth of crystals by surface adsorption, but both have been found to be less important in the absence of which stone occurrence could be more significant than citrate in clinical stone disease [19]. Macromolecular inhibitors include the urinary protein osteopontin (OPN), which is an aspartic acid-rich inhibitor that strongly inhibits the nucleation and growth of COM crystals in the presence of normal urine at concentrations of about 150 µg per milliliter and by stereochemically specific interactions of the carboxylate groups with calcium ions, pinning step advancement on particular crystal faces [20]. The most abundant urinary protein Tamm-Horsfall protein, on the other hand, affects the ability of CaOx crystals to aggregate based on the state of its polymerization, and glycosaminoglycans also interfere with crystal–cell surface interactions by occupying the anionic binding sites of the tubular epithelial membranes [21].
Crystal retention in the renal tubule is the final and decisive step in stone formation, which is also regulated by the crystal–cell adhesion. COM crystals interact quickly with the epithelial cells lining the distal tubules at anionic sites, beginning within seconds after the crystals come in contact with the cell surface, as shown in culture models with radio-labeled COM. The luminal surface of the oxalate-injured tubular cell displays significantly higher levels of hyaluronan, osteopontin and CD44, which mediates crystal adhesion, resulting in a vicious cycle of crystal deposition and oxidative injury. A schematic of the crystallization cascade is provided in Figure 1, and the main urinary promoters and inhibitors of CaOx crystallization and the mechanisms and modulation of these by phytochemicals are presented in Table 1 [22].
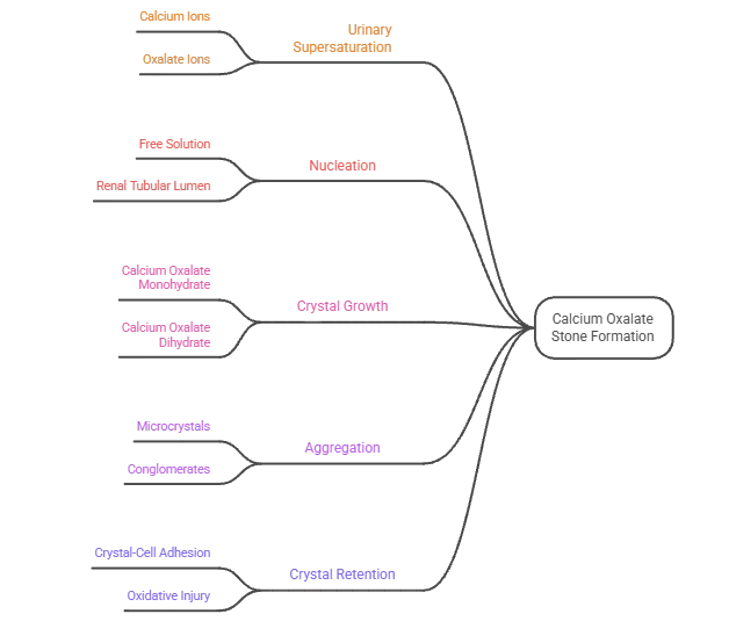
Figure 1. Pathophysiology of Calcium Oxalate Stone Formation
Table 1. Urinary Promoters and Inhibitors of Calcium Oxalate Crystallization: Physicochemical Roles, Normal Urinary Levels, and Documented Modulation by Plant-Derived Phytochemicals [23–27]
|
Factor |
Type |
Role in CaOx Crystallization |
Normal Urinary Level / Reference Range |
Modulation by Phytochemicals |
|
Calcium |
Promoter |
Provides ionic substrate for CaOx lattice formation; hypercalciuria drives supersaturation |
100–300 mg/day |
Flavonoids (e.g., quercetin) reduce urinary calcium in animal models via reduced intestinal absorption |
|
Oxalate |
Promoter |
Primary anion in CaOx crystal lattice; hyperoxaluria is the most common lithogenic metabolic abnormality |
20–45 mg/day |
Polyphenols reduce hepatic oxalate synthesis; plant extracts reduce urinary oxalate in ethylene glycol models |
|
Uric acid |
Promoter |
Promotes heterogeneous nucleation of CaOx by providing crystalline templates |
< 800 mg/day (men); < 750 mg/day (women) |
Alkaloids with xanthine oxidase inhibitory activity reduce urate production |
|
Sodium |
Promoter |
Increases urinary calcium excretion; competes with citrate reabsorption |
Variable; dietary-driven |
Limited direct phytochemical evidence; diuretic terpenes indirectly lower concentration |
|
Citrate |
Inhibitor |
Forms soluble calcium complexes; directly inhibits nucleation and aggregation via crystal face adsorption |
Mean ~640 mg/day; hypocitraturia < 320 mg/day |
Several flavonoids and organic acids in plants may raise urinary citrate levels in animal models |
|
Magnesium |
Inhibitor |
Forms soluble MgOx complexes; reduces free oxalate ion activity |
50–150 mg/day |
Magnesium-rich plant extracts raise urinary magnesium in hyperoxaluric rats |
|
Osteopontin (OPN) |
Dual regulator |
Inhibits COM nucleation and growth at normal urine levels; promotes crystal adhesion to CD44-expressing injured cells |
> 100 nmol/L in normal urine |
Kaempferol and related flavonoids downregulate OPN expression in crystal-injured HK-2 cells |
|
Tamm-Horsfall protein (THP) |
Inhibitor (physiological) |
Inhibits crystal aggregation in monomeric form; promotes aggregation when polymerized under pathological conditions |
Most abundant urinary protein; ~50 mg/day |
Indirect modulation via reduction of renal tubular injury; no direct phytochemical evidence reported |
|
Pyrophosphate |
Inhibitor |
Inhibits crystal growth through surface adsorption on COM crystal faces |
~50–100 μmol/day |
Not directly modulated by reported plant extracts |
|
Glycosaminoglycans |
Inhibitor |
Occupy anionic sites on tubular epithelial surfaces, reducing COM crystal adhesion |
Low nmol/L range |
Saponins in plant extracts may mimic glycosaminoglycan surface competition |
3. Role of Oxidative Stress and Inflammation in Crystal-Induced Renal Injury
Not all calcium is passively deposited as crystals in the renal tubular environment. When COM crystals are in contact with the apical surfaces of the renal tubular epithelial cells, they induce a cascade of intracellular events that result in a fundamental change of the cell's redox status, membrane integrity and inflammatory gene expression profile which paradoxically promotes further crystal adhesion and retention [28]. The knowledge of this biological amplification loop is crucial to understand if the role of phytochemical compounds as antioxidants and anti-inflammatory agents in anti-urolithiatic pharmacology makes sense [29].
When COM interacts with renal tubular epithelial cells, the intracellular NADPH oxidase activity is stimulated, in particular, through the p47phox subunit, which leads to an excess production of ROS such as superoxide anion, hydrogen peroxide and hydroxyl radical. These bursts of ROS exhaust oxidative stress defense mechanisms, such as superoxide dismutase, catalase, and glutathione peroxidase activities decrease significantly in cells exposed to crystals, and cause secondary markers of oxidative stress, such as increased malondialdehyde, a measure of membrane lipid peroxidation, and increased 8-hydroxydeoxyguanosine, a measure of oxidative modification of DNA [30]. Correspondingly, urinary concentrations of γ-glutamyl transpeptidase, angiotensin-converting enzyme, and N-acetyl-β-D-glucosaminidase are increased in CaOx stone formers compared to controls, indicating that tubulolithiasis is a systemic biochemical fact, and not just an experimental finding [31].
The oxidative injury to tubular cells leads to activation of 2 transcriptional axes central to crystal induced pathology. The first is NF-κB, which is activated and leads to the upregulation of expression of pro-inflammatory cytokines such as IL-1β, IL-6, TNF-α and the chemokine MCP-1, which promotes macrophage recruitment to sites of crystal deposition [32]. The latter and more recently characterized axis is the NLRP3 inflammasome, a multimeric complex in the cytosol that is activated upon ROS-dependent TXNIP signaling, followed by the activation of caspase-1, processing and secretion of mature IL-1β and IL-18. Using a hyperoxaluric rat model, it was concluded that there was a strong axis of ROS–TXNIP–NLRP3 signaling in the kidney with calcium oxalate stones, making the inflammasome a bona fide therapeutic target for stone disease. Critically, the inflammatory mediators produced by NLRP3 activation further promotes crystal aggregation, adhesion and retention within the tubules, thus exacerbating the initial oxidative insult which led to the activation of NLRP3 [33].
The Nrf2/antioxidant response element (ARE) pathway stands in opposition to this destructive cascade, and is the major endogenous cytoprotective system in the kidney against oxidative stress. In normal, basal conditions, Nrf2 is kept in the cytoplasm by its repressor, Keap1. The dissociation of Keap1 by oxidative stress results in the movement of Nrf2 to the nucleus where it regulates the expression of cytoprotective genes such as heme oxygenase-1, NAD(P)H quinone oxidoreductase 1, and catalytic and modulatory subunits of glutamate-cysteine ligase, which collectively raise the potential of the cells to combat oxidative stress [34]. Flavonoids from Lysimachia christinae were found to activate the Nrf2/ARE pathway in CaOx crystal challenged renal tubular epithelial cells and renal tissue, which indicates that botanical Nrf2 activators are able to activate this protective pathway in the context of nephrolithiasis. The pharmacologic profile of plant polyphenols and flavonoids (NFK, NLRP3, Nrf2) is one of convergence and underscores the mechanistic potential of multi-targeted chemoprevention approaches for stone prevention, compared to single-pathway pharmaceuticals [35].
4. Phytochemical Classes and Their Structural Basis for Antiurolithiatic Activity
The validity of plant-derived compounds as antiurolithiatic agents is based on a defensible chemical logic, a set of chemical features that account for the ability of certain compound classes to affect calcium oxalate crystallization, and the biology of renal injury. Before analysis of the individual plant data, it is important to review this structural pharmacognosy, to differentiate the activity that is likely mechanistically relevant from activity that may be non-specific or artifactual in vitro [36].
Flavonoids are the most extensively-studied subclass of polyphenols and have been the focus of the majority of studies conducted on the antiurolithiatic activity of polyphenols. The highly significant structural difference of polyphenols are the presence of several aromatic rings, each with hydroxyl groups; it is this structure, especially the two hydroxyl groups in the B ring of flavonoids (catechol), that gives them strong radical-scavenging properties via the capacity to donate electrons and chelate divalent metal ions, such as calcium [37]. In this context, the best studied flavonol is the multihydroxylated quercetin, with five hydroxyl groups arranged in the A, B and C rings. The presence of the 3-OH and 4-C=O groups on the C ring, along with the catechol on the B ring, provides a geometry that is ideally suited for bidentate ionic binding with calcium ions - directly relevant to the ionic environment of CaOx nucleation [38]. The study at different concentrations of quercetin (2.5 μM to 160 μM) showed that the inhibitory concentrations will be effective starting from the lowest concentration tested, indicating that activity is possible in physiologically relevant concentrations. The dual modulatory effects of quercetin, however, in the same experimental system, underscore the complexity of phytochemical-crystal interactions and the need to not rely on just one point of nucleation inhibition data but on a complete crystallization profile. Among the most mechanistically characterized antiurolithiatic flavonoids available in the literature, kaempferol is a structurally related flavonol, which lacked the 3′-hydroxyl group of the B ring, and showed the ability to inhibit CaOx crystal deposition in renal tubules and crystal adhesion to HK-2 cells, partly by inhibiting the aldose reductase/NOX2 signaling axis, with the accompanying reduction of OPN and CD44 expression in crystal-injured renal tissue [39].
The condensed proanthocyanidins and hydrolyzable gallotannins and ellagitannins possess antiurolithiatic activity mainly due to their surface-active properties. They have a high molecular weight, multiple hydroxyl groups and are anionic, which enables them to bind to the crystal surface and change the zeta potential of CaOx suspensions and lower the aggregation energy between particles [40]. Saponins are amphiphilic glycosides, with detergent properties, since they are composed of a lipophilic aglycone and polar sugar chains, which has been reported to disaggregate mucoproteins that form promoters of crystallization in urinary matrix of plants like Solanum xanthocarpum, phytochemical screening studies always reveal saponin rich fractions as responsible for the activity on antiurolithiatic effect. Berberine, an isoquinoline alkaloid with a planar polycyclic structure, is permanently charged and has documented XO inhibitory activity that is relevant to the reduction of urinary urate and anti-inflammatory activity through NF-κB suppression also provides a mechanistic contribution to reducing crystal induced injury [41]. The anti-urolithiatic importance of terpenoids is mostly due to anti-inflammatory COX pathway modulation and the diuretic activity (which increases urinary volume, diluting the solutes in urine that promote stone formation, and effectively flushing nascent crystals and preventing their aggregation and adhesion). The structural features, proposed mechanism(s) and representative plants of each major category of phytochemicals having antiurolithiatic properties are given in a systematic manner in Table 2, which also gives specific details of structural features, proposed mechanism(s) and representative plants for each phytochemical class for which no details are provided in the preceding prose [42].
Table 2. Major Phytochemical Classes Implicated in Calcium Oxalate Crystal Inhibition: Structural Features, Proposed Mechanisms, and Representative Compounds with Source Plants [43–45]
|
Phytochemical Class |
Representative Compounds |
Key Structural Feature Relevant to CaOx Inhibition |
Primary Antiurolithiatic Mechanism(s) |
Representative Source Plants |
|
Flavonols |
Quercetin, kaempferol, myricetin, fisetin |
Orthodihydroxy B-ring; 3-OH/4-carbonyl calcium chelation geometry; multiple phenolic –OH groups |
Calcium ion chelation reducing free ionic activity; CaOx nucleation inhibition; Nrf2/ARE activation; OPN/CD44 downregulation (kaempferol via AR/NOX2) |
Quercus spp., Allium cepa, Camellia sinensis, Aegle marmelos, Desmodium styracifolium |
|
Flavones |
Apigenin, luteolin, baicalin |
Planar 2-phenylchromone backbone; no 3-OH group distinguishes from flavonols |
Antioxidant ROS scavenging; NF-κB suppression; reduction of COM crystal adhesion to renal epithelium |
Petroselinum crispum, Scutellaria baicalensis, Matricaria chamomilla |
|
Flavan-3-ols (Catechins) |
Epigallocatechin gallate (EGCG), catechin, epicatechin |
Gallate ester at C-3; catechol and pyrogallol B-ring hydroxylation; no C2–C3 double bond |
SOD activity preservation; inhibition of crystal-induced lipid peroxidation; calcium complexation in urine |
Camellia sinensis (green tea), Vitis vinifera |
|
Hydrolyzable tannins |
Gallic acid, ellagic acid, punicalagin |
Polyhydroxybenzoyl ester linkages; high anionic charge density at physiological pH |
Crystal surface adsorption modifying zeta potential; aggregation inhibition; renal epithelial cytoprotection |
Punica granatum, Phyllanthus niruri, Terminalia chebula |
|
Condensed tannins (Proanthocyanidins) |
Procyanidin B2, proanthocyanidin A |
Polymeric flavan-3-ol units; high molecular weight with extensive –OH array |
Mucoprotein disaggregation reducing crystallization nucleation sites; crystal surface charge modification |
Vitis vinifera (grape seed), Vaccinium macrocarpon |
|
Saponins |
Diosgenin, solanine, sarsasapogenin glycosides |
Amphiphilic structure: lipophilic triterpenoid/steroidal aglycone + polar saccharide chain |
Disaggregation of urinary mucoproteins that serve as crystallization promoters; diuretic effects increasing urine volume |
Solanum xanthocarpum, Tribulus terrestris, Dioscorea spp. |
|
Isoquinoline alkaloids |
Berberine, palmatine |
Planar polycyclic quaternary nitrogen; permanently charged cationic structure |
Xanthine oxidase inhibition reducing urinary urate; NF-κB anti-inflammatory activity; modest crystal zeta potential modification |
Berberis vulgaris, Coptis chinensis, Mahonia aquifolium |
|
Pentacyclic triterpenoids |
Ursolic acid, oleanolic acid, betulinic acid |
Pentacyclic carbon skeleton with carboxyl and hydroxyl substituents |
COX-2 inhibition reducing prostaglandin-driven inflammatory response; diuretic activity lowering urinary solute concentration |
Rosmarinus officinalis, Ocimum sanctum, Malus domestica |
|
Phenolic acids |
Chlorogenic acid, rosmarinic acid, ferulic acid |
Hydroxycinnamic or hydroxybenzoic backbone; ester or amide linkages in plant matrix |
Free radical scavenging; modest calcium complexation through carboxylate groups; hepatic oxalate synthesis modulation |
Coffea arabica, Rosmarinus officinalis, Cichorium intybus |
5. In Vitro Evidence: Crystallization Assays, Cellular Models, and Key Findings
The three main experimental methods employed in vitro to assess the antiurolithiatic activity of plant extracts and isolated compounds involve the nucleation assay, the aggregation assay and the crystal growth assay, which are all carried out in turbidimetric system and the changes in optical density at 620 nm are used as an indicator of crystal growth kinetics [46]. In a typical protocol, CaCl₂ (usually between 4-8 mmol/L) is added to the sodium oxalate (usually between 0.5-1 mmol/L) solution in a buffered solution at 37°C and pH 5.5-6.5, which is similar to the pH and temperature of tubular fluid, and turbidity is monitored over time in the presence of the test compound and in its absence. A nucleation slope (SN) indicates the highest rate of the new particle formation, and an aggregation slope (SA) characterizes the aggregation processes of formed crystals. This acellular system allows for dose-dependent inhibition percentages to be calculated when compared to a drug-free control and, typically, a commercially available polyherbal reference, whose inhibitory activity is seen in the same manner but in a clinically contextualized manner [47].
There is extensive though inconsistent evidence from the published literature that comes from this method. The other extreme, a study comparing the nucleation inhibition of the aerial fraction of Aerva lanata containing phenols and flavonoids to that of the root fraction showed that the former has a significantly higher activity (71.01 ± 1.13%) than the latter (54.61 ± 2.30%) at the tested concentration, a result that is structurally coherent, since the aerial fraction is rich in phenols, whereas the root fraction is relatively low [48]. This role of the molecular weight and the carboxylate density has been clearly demonstrated with carboxymethylated Poria cocos polysaccharides, with a positive correlation of the degree of carboxymethylation to nucleation inhibition activity, conversion of COM to less adhesive COD polymorph, and decrease in the absolute value of crystal zeta potential; these results have direct structural analogy with the behaviour of carboxylate-rich polyphenols. Structure–activity correlation data for a panel of 19 molecules screened as inhibitors of CaOx crystal nucleation and growth further demonstrated that molecules with pKa <3.5, LogD at pH 6 <0, and HBI >0.1mmol/L are consistently more active at inhibiting CaOx crystal formation than molecules that do not meet these criteria, establishing a rational physicochemical profile for prioritizing compounds for antiurolithiatic screening campaigns [49].
The in vitro evidence-based data has a cellular dimension which is missing from acellular turbidimetric assays. The human proximal tubular epithelial cells (HK-2) exposed to oxalate or COM crystals is a model of crystal-induced renal injury and compounds may be tested for the ability to restore cell viability, normalize antioxidant enzyme activities, reduce ROS generation and suppress inflammatory gene expression. Kaempferol interfered with crystal deposition and adhesion in crystal-challenged HK-2 cells by modulating the AR/NOX2 pathway and it was found that kaempferol could significantly reverse the upregulation of adhesion molecules directly involved in amplifying crystal retention, such as OPN and CD44. The total flavonoids of Lysimachia christinae induced the Nrf2/ARE pathway in oxalate-challenged HK-2 cells, which, in turn, up-regulated the expression of the catalytic subunit of glutamate-cysteine ligase, and restored GSH levels, directly linking Nrf2 activation with crystal inhibition results [50]. The physical behaviour of injured cells versus intact cells is informative in itself, as oxidatively injured cells shift their crystal phase to COM (the more pathological polymorph) and induce COD production in normal HK-2 cells, a bridge between oxidative injury, crystal phase preference, and stone formation risk, which plant derived antioxidants could be gaining access to interrupt. The use of high concentrations is also repeatedly noted as an important drawback in the carefully designed studies in this field: Although as in the quercetin study, some concentrations are linked to human plasma levels, most exceed the urinary levels that could be obtained by standard dietary intake, suggesting that the biological plausibility of in vitro results needs to be tested against the pharmacokinetic reality of the human body before clinical relevance can be claimed. Mechanistic interactions between various classes of phytochemicals and CaOx crystal surfaces take place at three levels: suppression of nucleation, polymorphic conversion of COM to COD, and aggregation inhibition, which collectively define the crystallization modulation scope that can be achieved by using phytochemicals derived from plants [51].
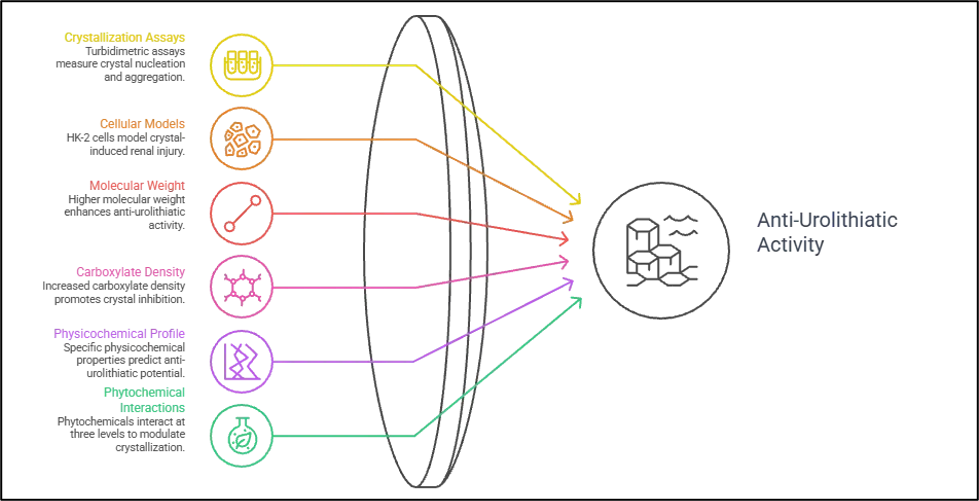
Figure 2. Mechanisms of Plant-Based Urolithiasis Inhibition.
6. In Vivo Validation and Transition to Preclinical Evidence
To interpret in vitro crystallization inhibition results to biologically relevant antiurolithiatic activity, validation in animal models that mimic clinical conditions for stone formation is necessary. The rat ethylene glycol–ammonium chloride model continues to be the most widely used preclinical model for this purpose [52]. Chronic intake of 0.75% ethylene glycol in drinking water results in chronic hyperoxaluria due to hepatic metabolism of ethylene glycol to glycolate and oxalate causing increased renal retention and excretion of calcium oxalate, which can be demonstrated by histopathological examination of renal tubular epithelia, and worsening renal function parameters such as elevated serum creatinine, serum blood urea nitrogen, urinary calcium, and urinary phosphate. The induction of oxidative stress in this model, i.e. increase of malondialdehyde and decrease of renal tissue activities of glutathione and catalase, is consistent with the previously reported cellular models, and this provides experimental coherence between the two experimental levels [53].
Therefore, a systematic review was carried out that included in vivo studies related to CaOx nephrolithiasis using medicinal plant extract preparation techniques and found that most of the plant organs, which were extracted by different techniques, exhibited strong antiurolithiatic activity and various mechanisms of action were reported, such as anti-oxidative and anti-inflammatory properties. Furthermore, a 2024 systematic review (from 2021 to 2023 publications) identified 64 studies that used PRISMA compliant methodology, which revealed that several plants and phytochemicals, including Alhagi maurorum, Aerva lanata, Dolichos biflorus, Cucumis melo, and quercetin, exhibited promise in reducing stone numbers, size, and formation in ethylene glycol models. The study on the effect of the extract of the bark of Ficus racemosa on kidney structure and function in ethylene glycol induced diabetic rats was a representative study in this category because the level of glutathione and catalase were significantly reduced and malondialdehyde was significantly increased in the kidney, compared to normal control, on day 14 and 28 of ethylene glycol induction, whereas the treatment groups showed significant increase in the level of glutathione, decrease in malondialdehyde and histopathology of the kidney, which showed reduced calcium oxalate crystal mediated cellular damage. In a 15-day ethylene glycol model, the polyphenol-rich fraction of A. spinosa corrected the elevated urinary pH and protein, restored the decreased magnesium and reversed the renal and hepatic tissue lesions, while molecular docking analysis showed that the polyphenols with the highest content exhibited high binding affinity with the molecular targets involved in urolithiasis, highlighting the growing use of in silico techniques in preclinical studies for the treatment of urolithiasis [54].
The major drawback of this in vivo evidence base is the predominant use of the ethylene glycol model with pharmacological properties of very high oxalate loads and rapid, heavy crystal deposition, which are not representative of slower, metabolically mediated crystallization conditions of human idiopathic nephrolithiasis. The model also fails to take into account differences between the effects of a test compound on the actual inhibition of crystallization and secondary effects that occur due to changes in urine volume, urine pH or oxalate absorption from the gut. In addition, there are no clear conclusions that can be drawn from in vivo data to structure–activity relationships since there is no consistency in the characterization of the extracts, and it is not clear what fraction of the phytochemicals is responsible for the observed activity. Clinical trial data in this area is limited, but a randomized controlled trial that found a phytochemical-rich lime-based preparation reduced the recurrence of CaOx urolithiasis by 76% after 24 months, is one of the more compelling pieces of clinical trial evidence, although the authors noted that results need to be confirmed in larger and more diverse patient groups before the clinical implications can be drawn [55].
7. Bioavailability, Metabolic Transformation, and Renal Delivery Considerations
The therapeutic credibility of any plant compound, whether it is an antiurolithiatic or any other compound, will ultimately depend upon whether adequate level reaches the targeted biological compartment (in the case of antiurolithiatic, renal tubular fluid and tubular epithelium) after oral administration under physiologically realistic conditions. This pharmacokinetic dimension is the one which has been least studied in the literature of the antiurolithiatic phytochemicals, and is the most crucial gap that has to be translated between the in vitro and in vivo studies of high pharmacological significance and the clinical trial results that can produce evidence-based interventions.
The predominant form in plant matrices is the flavonoid glycoside and it undergoes sequential hydrolysis during gastrointestinal transit. Quercetin-3-glucoside and related glucose conjugates undergo hydrolysis at the brush border of enterocytes to release the aglycone, which in turn is passively absorbed. Rutin and other rhamnosylated flavonoids must survive to the colon where they are hydrolyzed by α-rhamnosidases secreted by Bifidobacterium species to release the aglycone for colonic absorption. After absorption, quercetin aglycone is highly conjugated, mainly to glucuronides and sulfates, in the small intestinal epithelium and in the liver, leading to plasma pools which are dominated by quercetin glucuronides and sulfates instead of the aglycone. This first pass metabolism is consistently reported to result in poor oral bioavailability of quercetin, of which Cmax in plasma after a 50 mg dose is around 4 μmol/L for metabolites, and about 38% of the absorbed dose is excreted in urine, but the amount excreted as intact aglycone or biologically active metabolites, with the remaining CaOx inhibitory activity, is markedly less than the total conjugated urinary flavonoid load. The ellagitannins found in pomegranate and berries are poorly absorbed in the small intestines and make their way to the colon where bacterial biotransformation produces urolithins, metabolites that are found in kidney tissue within 1 to 6 hours after oral urolithin A intake and have anti-inflammatory and antioxidant activity different from the ellagitannin. In order to provide researchers with a reference that brings together published data for oral bioavailability, Cmax, plasma half-life, urinary excretion, and known metabolites, the pharmacokinetic properties of some of the most studied flavonoids and phenolic acids are given in Table 3 [56].
The implication of these pharmacokinetically derived facts is that the inhibitory concentrations observed in vitro may not be possible to obtain in the urinary compartment with standard oral administration of unformulated plant extracts. The use of lipid-based nanocarrier systems like liposomes, nanostructured lipid carriers (NLC), and solid-lipid nanoparticles (SLC) has been found to significantly enhance the oral bioavailability and antioxidant effects of quercetin by bypassing metabolism of the enterocytes' brush border, and by boosting lymphatic uptake. Phytosome formulations have also shown to have a higher membrane permeability and less metabolic degradation by intestinal enzymes; for example, in fecal incubation in vitro the percentage of quercetin degraded by the gut flora was lower than that of the formulation with a phytosome, suggesting that more of the active compound might be available for absorption. The use of nanotechnology platforms to deliver nanocrystals of quercetin to the cytoplasm of renal cells has been demonstrated to improve its solubility and antioxidant activity, creating a pharmacologically achievable goal when appropriate nanotechnology platforms are used. A representative flavonoid - ingestion to biotransformation to tubular secretion to urinary accumulation - is shown conceptually in the gut–liver–kidney disposition axis [57].
Table 3. Pharmacokinetic Parameters and Urinary Disposition of Key Antiurolithiatic Phytochemicals: Oral Bioavailability, Plasma Cmax, Metabolic Pathways, Active Urinary Metabolites, and Formulation Enhancement Strategies [58]
|
Compound |
Class |
Oral Bio-availability (%) |
Reported Plasma Cmax (typical dietary/ supplemental dose) |
Plasma t½ |
Urinary Excretion (% of absorbed dose) |
Key Active Metabolites with Anti-urolithiatic Relevance |
Formulation Strategy Improving Absorption |
|
Quercetin |
Flavonol |
Low (variable, <10% as aglycone; higher as glucoside) |
~4 μmol/L (metabolites, 50 mg dose) |
11–28 h (conjugates) |
~38% (as conjugates) |
Quercetin-3-glucuronide; isorhamnetin (methylated metabolite); 3,4-dihydroxyphenylacetic acid |
Phytosome, solid-lipid nanoparticles, nanostructured lipid carriers |
|
Kaempferol |
Flavonol |
Low; similar profile to quercetin with less studied glycoside fraction |
~1–2 μmol/L (estimated) |
~6–24 h (conjugates) |
Low; primarily as glucuronide conjugates |
Kaempferol glucuronides; 4-hydroxyphenylacetic acid (colonic catabolite) |
Co-delivery with phospholipids; cyclodextrin inclusion |
|
Gallic acid |
Phenolic acid |
Moderate (~70%) as free acid; lower as gallotannin ester |
~2–5 μmol/L |
Short (~1.6 h) |
~35–40% |
Pyrogallol (bacterial catabolite); 4-O-methylgallic acid |
Generally good absorption as free acid; formulation less critical |
|
Ellagic acid / Ellagitannins |
Phenolic acid / Hydrolyzable tannin |
Very low as intact molecule; high colonic biotransformation |
Ellagic acid Cmax <100 nmol/L; urolithins higher |
Ellagic acid t½ ~0.9 h; urolithin t½ much longer |
Urolithins (A, B, C) detected in urine and kidney tissue |
Urolithin A — anti-inflammatory, antioxidant; renal tissue accumulation confirmed |
Formulation less relevant; microbial conversion is the primary activation step |
|
Apigenin |
Flavone |
Low; glycoside (apiin) hydrolyzed by colonic bacteria |
~1 μmol/L (estimated) |
~6–12 h |
Low; as flavone glucuronides |
Luteolin (minor oxidative product); phenolic acid catabolites |
Nanoemulsion, self-nanoemulsifying delivery systems |
|
Berberine |
Isoquinoline alkaloid |
Very low (~5%) due to P-glycoprotein efflux and poor membrane permeability |
~0.5–1 μmol/L (500 mg dose) |
~10–30 h |
<5%; extensive hepatic metabolism |
Berberrubine; jatrorrhizine; demethylene-berberine |
Nanoparticle encapsulation; phospholipid complexation significantly improves Cmax |
|
Curcumin |
Diarylheptanoid |
Extremely low (<1%) due to rapid metabolism and poor solubility |
Negligible as free compound |
Very short (<1 h) |
Negligible as free compound; tetrahydrocurcumin detected in urine |
Tetra-hydrocurcumin; curcumin glucuronides; curcumin sulfates |
Critical formulation need: self-nanoemulsifying systems, polymeric nanoparticles, and phytosomes dramatically improve Cmax |
8. Limitations of Current Evidence and Translational Gaps
The body of evidence reviewed in the preceding sections is broadly substantiated and mechanistically informative, but is structurally limited, in the manner in which antiurolithiatic phytochemical research has been designed and conducted in the majority of published studies. It is not a deprecation of the promise of the field to acknowledge these limitations but a necessary requirement for designing studies that will answer the question of whether the promise of the field can be realized clinically. The first and most obvious limitation is that there are differences between the in vitro inhibitory activity and attainable urinary levels in humans. Most spectrophotometric crystallization assay studies are performed at a concentration of 0.1–5.0 mg/mL or equivalent molar dosage, which are far higher than what can be reliably provided to the urinary compartment by the oral administration of standardized plant extracts or isolated compounds, especially in human subjects where bioavailability is poor for compounds like curcumin and berberine. Whereas the few studies that have tried to correlate assay concentrations with human blood or urine levels, like the quercetin study (2.5-160 μM), are remarkable for being rare exceptions. This disparity results in a significant percentage of positive in vitro data in the antiurolithiatic literature that correspond to a real effect in the physicochem domain but cannot be reached without formulation intervention in the pharmacokinet domain [59].
The second constraint is the lack of standardization of plant extracts in the experimental studies. The antiurolithiatic activity described for an extract is the sum total of the variations in qualitative and quantitative phytochemical profile that can occur between extracts of the same plant, but different extraction solvents, plant parts employed, geographic origin, harvest time, and processing method. Chemical fingerprinting, preferably by HR-LCMS, and reporting of the principal active constituent(s) and their concentration is required to compare activity data between studies and to confidently identify a compound or compounds that are responsible for the observed inhibition in order to make mechanistic claims. The inability to standardise across the heterogeneous studies is evident in the systematic review literature and is the reason why inferential power of meta-analytical approaches in this domain is limited.
The third constraint is the lack of a sex-specific analysis in the majority of pre-clinical studies. An epidemiological sex difference is apparent, with calcium oxalate stones being formed around twice as often in men than in women, and urinary citrate levels being lower in women than in men, thus setting up a different inhibitory environment. Sex hormones affect oxalate metabolism, calcium handling in the kidney and excretion of inhibitors of calcium handling in the urine in a biologically significant manner that is usually not accounted for in model studies using rats, which are typically male. This lack of generalizability of preclinical results to the broader patient population is a limitation.
A fourth issue is the fact that in the patients who will be actually taking conventional antiurolithiatic medicines, attention is limited to herb–drug interactions. High polyphenol loads can interact with cytochrome P450 enzymes (e.g. CYP3A4 and CYP2C9 are inhibited by quercetin and will change the plasma concentration of some drugs, including thiazide diuretics and allopurinol, which are used to treat nephrolithiasis; this interaction has not been studied in the context of nephrolithiasis pharmacotherapy). Lastly, there is not enough and not consistent enough clinical trial data to recommend at the guideline level. The limited randomized controlled trial evidence is directionally supportive, but consistently reports low quality of evidence, small sample sizes, and insufficient reporting of baseline urinary biochemistry to guide clinical practice [60].
9. Future Perspectives
The area of research of immediate relevance to the above mentioned translational bottlenecks is those that can provide the necessary methodological and clinical infrastructure to take phytochemical antiurolithiatic drugs into the evidence-based therapeutic recommendations in clinic. Network pharmacology and molecular docking have proven to be powerful front-end tools to define the multi-target pharmacological spectrum of complex plant extracts prior to investing in wet laboratory experiments. When applied to Choerospondias axillaris, quercetin, kaempferol, catechin, β-sitosterol and naringenin have been identified as major active compounds, AKT1, IL-6, TNF, TP53 and IL-1β have been emerged as main molecular targets and NF-κB and MAPK have been identified as the key signaling pathways involved by the application of PPI network analysis and KEGG pathway enrichment analysis. Importantly, these computational predictions were confirmed in experiment: experiments were performed in the 2-D CaOx agar gel system, where the extract at high concentrations exhibited a shift in crystal polymorph, from COH to COD, which is the polymorph that is associated with less epithelial adhesion and lower pathological risk. Incorporating this integrated computational-experimental approach systematically, across the most promising plant families that exhibit high densities of compounds that satisfy the criteria for good CaOx inhibition (pKa, LogD and hydrogen-bond) would significantly speed up the discovery of clinical lead compounds in antiurolithiatic medicinal chemistry. The case study of ellagic acid in Phyllanthus niruri is a typical success of such an approach, which was predicted using a network pharmacology approach to be targeting the proteins SQLE, SCD and HMGCS1 involved in CaOx-induced renal injury, and subsequently validated in vitro and in vivo [61].
The formulation challenge described in Section 7 requires a particular approach to research, which is the development of a delivery system to optimize the systemic bioavailability in a more conventional sense, but rather one that maximizes urinary excretion of an active or pro-active compound. There were no published studies that specifically characterized urinary pharmacokinetics of n LDs antiurolithiatic flavonoids (the urinary Cmax and the course of urinary inhibitory activity) as the main pharmacokinetic endpoint although the proof of concept of the oral absorption of these flavonoids by LDs with lipids and phytosomes was demonstrated. Such studies would bring about a pharmacokinetic–pharmacodynamic linkage for which the field is currently lacking, by combining the nano-enabled delivery with the validated in vitro crystallization assays performed at urine concentrations achieved in vivo. With proven regulatory expertise in nutraceutical and pharmaceutical applications, self-nanoemulsifying drug delivery systems have significantly enhanced the oral bioavailability of poorly soluble polyphenols such as curcumin and thus provide a scalable and practical system for this purpose.
Standardization of the conditions of the in vitro antiurolithiatic assay, such as crystallization buffer composition, pH, ionic strength, temperature, incubation time, and turbidity measurement parameters, within research groups would significantly enhance the comparability of the data reported from various laboratories, which would allow meaningful meta-analyses of concentration–response relationships. It is also critical that the use of extract characterization by HR-LCMS is established as a minimum reporting standard, which correlates observed activity to a defined phytochemical fingerprint, instead of a crude extract mass. From a clinical perspective, the design of Phase II randomized controlled trials of standardized, characterized phytochemical preparations in well-phenotyped cohorts of recurrent CaOx stone-formers with 24-hour urine biochemistry, stone recurrence imaging and urinary biomarkers of oxidative stress as co-primary outcomes represents the most straightforward pathway to guideline-relevant evidence. The promising preliminary results of clinical trials with clinical formulations that are rich in phytochemicals and have been shown to reduce the recurrence to a significant degree support the need to pursue such trials as a research priority [62].
CONCLUSION
The plant-based bioactive compounds are not only a mechanistically coherent approach to calcium oxalate stone disease, but they target multiple pathways simultaneously, such as the ability to inhibit the crystallization process, protect against oxidative stress, and suppress inflammation all of which are not provided by the pharmaceutical agents available today that target just one pathway. Based on the reviewed evidence, polyphenols, flavonoids, tannins, saponins, alkaloids and terpenoids have proven to possess structurally explicable antiurolithiatic effects backed by in vitro, preclinical and early clinical evidence. Gaps identified such as the concentration achievability problem, lack of extract standardization, lack of sex differentiated preclinical design, and lack of clinical trial evidence are research design issues, not biological in nature. Whether plant derived crystallization modulators can be incorporated in to evidence-based components of nephrolithiasis prevention will be determined by addressing them with HR-LCMS guided standardization, nano-enabled renal delivery strategies and adequately powered clinical trials.
REFERENCES


Aradhana Kangare, Vilas Ghawate, Varsha Jadhav, Snehal Rahate, Jayashri Shejul, Ethnopharmacological and Phytochemical Basis of Medicinal Plants in Urolithiasis: From Traditional Use to Molecular Mechanisms, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 6, 3083-3104. https://doi.org/10.5281/zenodo.20660424
 10.5281/zenodo.20660424
10.5281/zenodo.20660424