
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
Department of Biotechnology Jayoti Vidyapeeth Women’s University, Jaipur.
Salinization remains one of the most prevalent abiotic limiting factors for vegetable production globally and vegetables are especially sensitive due to their shallow root system and high transpiration rates in leaves, as in the case of spinach (Spinacia oleracea L.). This study was designed to assess the effect of green synthesized zinc oxide nanoparticles (ZnO-NPs) using sustainable route from Azadirachta indica (neem) leaf extract on NaCl induced damage in spinach. A 100% factorial completely randomized design was employed with two salinity levels (0 and 100 mM NaCl) and five levels of foliar application of ZnO-NP (0, 25, 50, 75 and 100 mg L-1) replicated three times under greenhouse conditions (25 ± 2 °C, 60 ± 5 %RH, 16 h photoperiod). Planting was performed and salinity was applied step-wise from day 21 and foliar sprays were done at 7-day intervals for 49 days after sowing. Salinity alone (S) significantly reduced plant height (37 %), leaf area (44 %), fresh (42 %) and dry (41 %) biomass, the SPAD index and total chlorophyll, while causing an increase of electrolyte leakage (EL) by nearly 3.4-fold, an increase of shoot Na? by 9.2-fold, and a decrease of K?/Na? ratio from 15.82 to 0.91. All these deleterious effects were significantly reduced in a dose dependent manner with the maximum dose (S+Z?: 75 mg L?¹) consistently being the most effective. Salinity stress resulted in a recovery of plant height in S+Z? (to ~88 % of the non-stressed control), and of leaf area (to ~90 %), fresh and dry biomass (to ~91 % and ~89 % of the non-stressed control, respectively), and a restoration of Fv/Fm (0.784) and of RWC (82.2 %), while EL was reduced to 18.53 %, shoot Na? content was lowered by ~59 % and shoot K?/Na? ratio was increased by more than 4-fold of the non-stressed control. Slightly reduced protection was observed at the 100 mg L?¹ dose, suggesting a saturation or mild toxicity level. Neem mediated ZnO-NPs are thus a sustainable low input approach to ensure the spinach productivity under salt affected horticulture and 75 mg L-1 is the agronomically optimum foliar dose.
Salinization of soil is a significant and growing soil degradation problem that currently affects over 1.38 billion hectares of land worldwide and is estimated to comprise 20–33 % of the world's irrigated farmland (FAO, 2021; Hassani et al., 2021). Secondary salinization is the result of climate change, sea-level rise, use of poor-quality irrigation water, excessive fertilizer uses and inadequate drainage, and is accelerating at a global scale, such that nearly half of all arable land is projected to experience secondary salinization by 2050 (Shahid et al., 2018; Hassani et al., 2021). The adverse effects of elevated soil Na⁺ and Cl⁻ on plant's physiology proceed sequentially: Osmotic phase (inhibiting water uptake); Ionic phase (resulting from Na⁺/K⁺ imbalance and specific ion toxicity); Secondary oxidative phase (excess accumulation of reactive oxygen species (ROS)) (Munns & Tester, 2008; Isayenkov & Maathuis, 2019).
Leafy vegetables (cultivated for fresh foliar biomass) are especially sensitive to salinity damage. Spinach (Spinacia oleracea L.) is an important leafy crop with high levels of iron, folate, β-carotene, vitamin C and vitamin K and antioxidant phytochemicals (Roberts and Moreau, 2016). Spinach is moderately tolerant to salinity than other Amaranthaceae, salinity levels above 80-100 mM significantly reduce the leaf number, fresh weight, chlorophyll, photosynthetic efficiency and nutrient quality (Ors & Suarez, 2017; Saleem et al., 2021). Due to the direct correlation between leaf surface area/water relations and marketable yield of spinach, moderate salinity will result in significant economic loss (Ors & Suarez, 2017).
Traditional practices to control salt affected soils such as gypsum amendments, leaching with quality irrigation water, drainage installation and cultivation of salt-tolerant cultivars are capital intensive and time consuming, and sometimes impractical for smallholder vegetable producers (Qadir et al., 2014; Shahid et al., 2018). Stress amelioration by exogenous application of osmoprotectants, plant growth regulators and macromolecules like Ca²⁺ or K⁺ usually requires multiple applications of soil drenching and provides inconsistent and low recovery (Shahid et al., 2018). So, there is an urgent need of sustainable, low input and field deployable technologies to protect the leafy vegetables grown on marginal saline land.
Recently, plant nanobionics, particularly metal-oxide nanoparticles applied as foliar spray, has been shown to be a promising approach for increasing plant resilience to abiotic stress (Khan et al., 2017; Singh et al., 2022). Among the most studied are the zinc oxide nanoparticles (ZnO-NPs) which play a central role in over 300 enzymes, in carbonic anhydrase, in Cu/Zn superoxide dismutase, in indole-3-acetic acid biosynthesis, in chlorophyll biosynthesis and in stabilizing membrane structure (Cakmak, 2000; Sturikova et al., 2018). In cereals, salinity, drought or heavy metal stress, exposure to which has improved growth, photosynthesis, antioxidant defence and ion balance, several reports have been published showing the improvements in growth traits of cereals due to exposure of ZnO-NPs under salinity, drought and heavy metal stress (Faizan et al., 2018; Rizwan et al., 2019; Zafar et al., 2016). Similarly, for legumes, improved growth traits under salinity stress and zinc have been reported with exposure to ZnO-NPs (Venkatachalam et al., 2017), while in tomato, growth enhancement under heavy metal stress has been shown (Faizan et al., 2021).
However, there are two key challenges that are still not addressed. First, the majority of the ZnO-NPs studied till now have been synthesized using chemical or physical methods using toxic reductants, high-temperature calcination and significant energy consumption, which imposes sustainability and biosafety concerns (Iravani, 2011; Singh et al., 2018). Green synthesis using plant extracts (particularly Azadirachta indica or neem) is an environmentally friendly single pot process where various secondary metabolites, such as nimbin, azadirachtin, polyphenols and terpenoids not only reduce the Zn²⁺ ions but also cap the resulting nanoparticles thereby forming nanoparticles with enhanced biological activity and colloidal stability (Elumalai et al., 2015; Bhuyan et al., 2015; Madan et al., 2016). Second, research on the dose-dependent protective effect of green-synthesized ZnO-NPs in spinach under salinity is limited while spinach is among the most widely cultivated leafy vegetables in salt-affected soils in South and Southeast Asian countries.
Therefore, the objectives of the present study was to synthesize ZnO-NPs by using an aqueous neem leaf extract, investigate the individual and combined effects of foliar application of ZnO-NPs (25, 50, 75 and 100 mg L⁻¹) and salt stress (100 mM NaCl) on growth (plant height, leaf area, fresh and dry biomass), photosynthetic performance (SPAD index, total chlorophyll, Fv/Fm), water relations (RWC), membrane integrity (electrolyte leakage) and ionic homeostasis (shoot Na⁺, K⁺ and K⁺/Na⁺ ratio) of Spinacia oleracea, and to find the optimum foliar dose for ameliorating salinity in spinach. We suspected that foliar application of the ZnO-NPs will be dose dependent, such that an intermediate rate (ca. 75 mg L⁻¹) would be optimal for reducing the salt-induced damage, while at the same time increasing the enzymatic functions mediated by Zn, photosynthetic efficiency, membrane stability and K⁺/Na⁺ homeostasis.
2. MATERIALS AND METHODS
2.1 Plant material and growth conditions
Certified seeds of Spinacia oleracea L. cultivars of 'All Season' were sourced from a registered seed supplier. After visual sorting to eliminate damaged and discoloured seeds, the seeds were surface-sterilized for 10 min in 1 % (v/v) sodium hypochlorite solution, washed five times in sterile distilled water and imbibed in distilled water for 12 h to ensure the synchronization of germination (Rodrigues et al., 2020). The soil in the field was autoclaved for 45 minutes on each of two consecutive days and mixed with coarse river sand and perlite in a ratio of 3:1:1 (v/v/v) to form a uniform medium (pH 6.8 ± 0.2; EC < 0.4 dS m⁻¹). Five seeds were sown at 1 cm depth in pots (diameter 20 cm and height 18 cm; medium holding capacity ~2 kg). Plants were grown in a greenhouse at 25 ± 2 °C, 60 ± 5 %RH and 16 h photoperiod with twice-weekly irrigation by both the tap water and half-strength Hoagland's nutrient solution until the start of the treatments.
2.2 Green synthesis of ZnO-NPs
Healthy A. indica leaves were collected and washed Three times (tap water, 0.1 % Tween-80 and distilled water), shade dried and grinded through 60-mesh sieve. The leaf powder was extracted using 1 L of ultrapure water at 80 °C for 60 min with continuous stirring (300 rpm) and filtered through muslin cloth and Whatman No. 1 paper and used within 2 h. The preparation of ZnO-NPs was performed by mixing 0.1 M zinc acetate dihydrate solution with neem extract (2:1, v/v) at 50 °C, and after 2 h, when the precipitation began, the mixture was heated to 80 °C for 2 h while it was stirred magnetically (400 rpm) following Elumalai et al. (2015). The resultant pale-white suspension was centrifuged at 10,000 rpm for 40 min, washed 3 times with ultrapure water (with intermittent probe-sonication) and re-suspended in absolute ethanol before drying for 40 min at 60 °C until constant weight. The dried nanopowder was kept in amber glass vials with silica gel until it was needed.
2.3 Salinity induction and ZnO-NP foliar treatment
Treatments were made 21 days after sowing (DAS) when seedlings reached four true leaves. To prevent osmotic shock, the various concentrations of salt (NaCl) were added in steps: 25 mM on days 1 to 2, 50 mM on days 3 to 4, 75 mM on days 5 to 6, and 100 mM from day 7 onwards (200 mL of salt solution per pot every 2 days). The conductivity of the 100 mM NaCl irrigation solution was 9.8 dS m⁻¹. Freshly prepared foliar suspensions of ZnO-NP (25, 50, 75 and 100 mg L⁻¹), were prepared by dispersing the dried nanopowder in distilled water, with 0.05 % (v/v) Tween-80, immediately prior to spraying, and subjected to probe-sonication for 10 min (40 % amplitude). Calibrated fine-mist hand atomizers were used to apply sprays every seven days (21-49 DAS) on the adaxial and abaxial leaf surfaces. Plants in control treatments were treated with the same amount of 0.05 % Tween-80 solution in distilled water.
2.4 Experimental design
The experiment was designed as a completely randomized design (CRD) with ten treatment combinations: Control (C), Salt stress (S, 100 mM NaCl), Z₁–Z₄ (25, 50, 75 and 100 mg L⁻¹ ZnO-NPs without salt) and S+Z₁–S+Z₄ (the same ZnO-NP doses with 100 mM NaCl). Triplicate treatments were used for each treatment.
2.5 Growth parameters
Harvesting was done at 49 DAS between 08:00 and 10:00 h to reduce the diurnal variation. Plant height (cm) was measured from soil surface to the longest tip of leaves. Total leaf area per plant (cm²) was determined with a LI-COR LI-3100C leaf area meter. Fresh leaf weight was harvested at once while dry leaf weight was harvested after the leaves were oven dried at 70 °C until constant weight.
2.6 Photosynthetic parameters
The chlorophyll content of leaves was non-destructively estimated with a Konica Minolta SPAD-502 Plus meter (three readings were taken on base, middle and tip of each leaf and averaged) and calculated as total chlorophyll (mg/g FW) using a calibration equation of Markwell et al. (1995). The chlorophyll a fluorescence was determined after 30 min of dark adaptation, using a Heinz Walz Mini-PAM fluorometer, where the F₀ was measured under a measuring light of 0.05 µmol m⁻² s⁻¹ and the Fm was measured under a saturating pulse (8000 µmol m⁻² s⁻¹, 0.8 s). The Fv/Fm ratio was calculated as: (Fm − F₀)/Fm = maximum quantum yield of photosystem II.
2.7 Water relations and membrane integrity
Relative water content (RWC) was calculated according to Barrs and Weatherley (1962) as follows:
RWC (%) = (FW − DW)/(TW − DW) × 100,
where the fresh weight (FW), turgid weight (TW) and dry weight (DW) of the leaf discs were after 4 h floating in distilled water and after 48 h at 70 °C, respectively
Electrolyte leakage (EL) was measured as described by Lutts et al. (1996) – 6 leaf disks (~100 mg FW) were placed in 10 mL of deionized water and shaken on a rotary shaker for 2 h at room temperature, followed by autoclaving for 20 min at 121 °C and measuring the conductivity after cooling (EC₂), which was subtracted from the initial conductivity (EC₁).
EL (%) = (EC₁/EC₂) × 100.
2.8 Shoot Na⁺, K⁺ and K⁺/Na⁺ ratio
The dried shoot tissue (0.2 g) was digested in 10 mL of HNO₃ + 2 mL of 30 % H₂O₂ at 80 °C for 6 h and then made up to 25 mL with 2 % HNO₃ and analysed by flame atomic absorption spectroscopy (AA) at a wavelength of 766.5 nm (Na⁺) and 589 nm (K⁺) against certified multi-element standards. Elemental concentrations were used to calculate the shoot K⁺/Na⁺ ratio, which is a measure of ionic homeostasis (Munns & Tester, 2008).
2.9 Statistical analysis
The two-way analysis of variance (ANOVA) was used for all data, which were tested for the 2 × 5 factorial CRD with IBM SPSS Statistics 29.0. Treatment means (n = 3) separated by Tukey's HSD test at p < 0.05 are presented as mean ± standard error (SE). Significance levels are reported as * p < 0.05, ** p < 0.01 and *** p < 0.001.
3. RESULTS
3.1 Growth response: plant height, leaf area and biomass
Salinity (100 mM NaCl) significantly lowered all growth parameters of spinach compared to the non-stressed control (Tukey HSD, p < 0.05; Figs. 1–3, Table-1). Plant height decreased from 28.33 cm in C to 17.86 cm in S, a 36.9 % reduction. Non-stressed plant foliar application of ZnO-NPs showed a slight stimulatory effect on the growth parameter (height; Z₁–Z₄; 29.25–30.66 cm), while the most noticeable response was obtained in the case of a combination of ZnO-NPs and salinity. Salt stressed plants demonstrated a gradual increase in height with increasing dose of salt stress (S+Z₁ 20.51 cm; Tukey 'd' to S+Z₃ 25.01 cm; Tukey 'c'), having recovered about ≈88 % of the control height. The 100 mg L⁻¹ dose (S+Z₄, 23.93 cm) provided slightly less protection than 75 mg L⁻¹, indicating a saturation/mild-toxicity threshold (ANOVA F = 95.34; p = 1.91 × 10⁻¹⁴).
Total leaf area decreased from 93.71 cm² (C) to 52.20 cm² in S (−44.3 %; Fig. 2). Among non-stressed plants, the 50 and 75 mg L⁻¹ doses produced the largest leaf area (Z₂ = 101.94 cm²; Z₃ = 101.90 cm²; Tukey 'a'). Under salinity, leaf area recovery followed the order S+Z₃ > S+Z₂ ≈ S+Z₄ > S+Z₁: S+Z₃ reached 84.19 cm², equivalent to 89.8 % of the control, while S+Z₁ (64.25 cm²) and S+Z₄ (79.37 cm²) recovered to 68.6 % and 84.7 %, respectively (F = 64.86; p = 7.83 × 10⁻¹³).
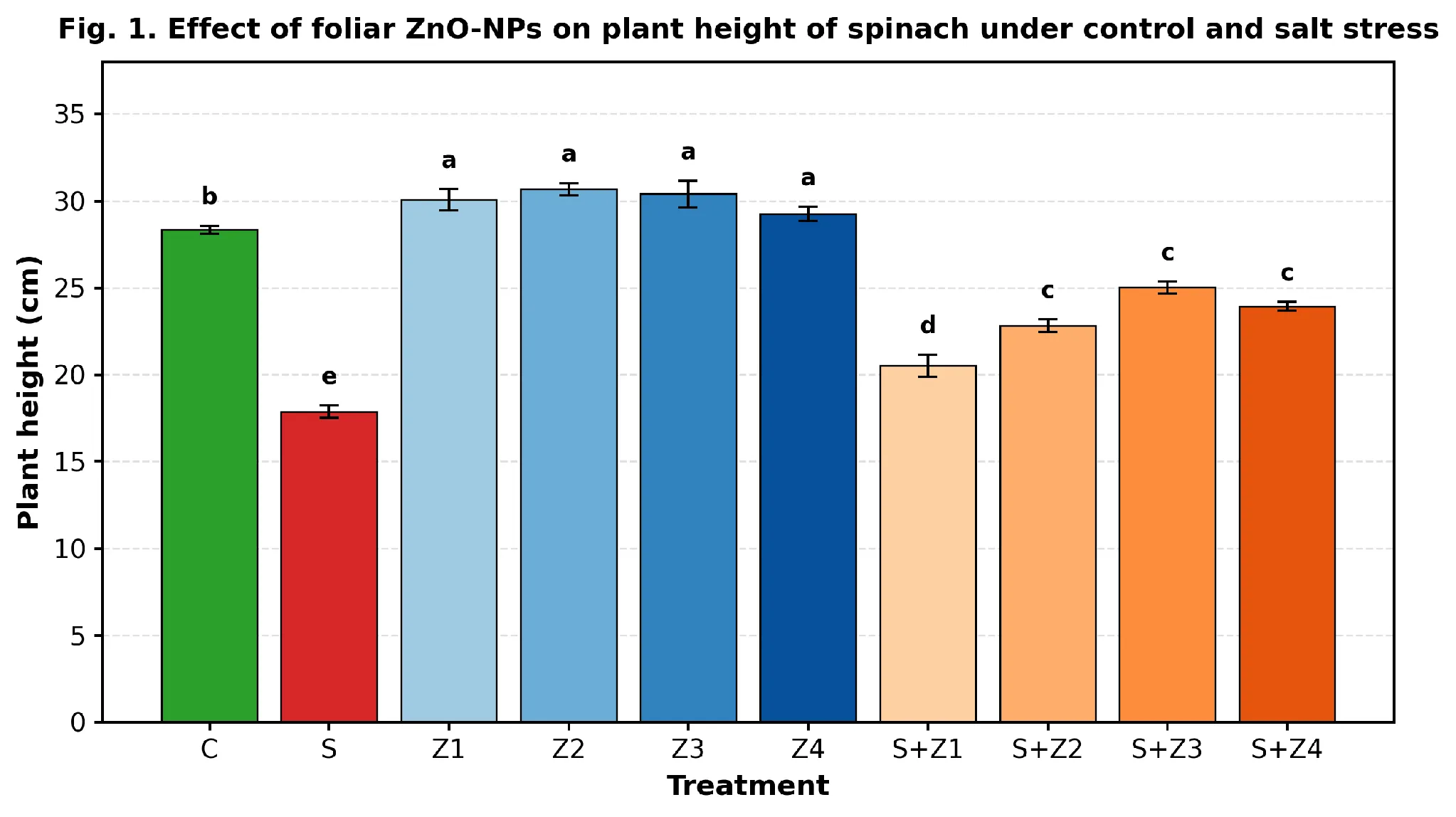
Fig. 1. Effect of foliar ZnO-NP application on plant height (cm) of Spinacia oleracea under non-stressed (C, Z₁–Z₄) and 100 mM NaCl-stressed (S, S+Z₁–S+Z₄) conditions. Bars represent means ± SE (n = 3). Different letters above bars indicate statistically significant differences (Tukey HSD, p < 0.05).

Fig. 2. Effect of foliar ZnO-NP application on total leaf area per plant (cm²) of spinach under control and salt-stress conditions. Bars represent means ± SE (n = 3); different letters indicate significant differences at p < 0.05 (Tukey HSD).
Salinity reduced fresh leaf weight from 8.647 to 4.991 g plant⁻¹ (−42.3 %) and dry weight from 1.070 to 0.632 g plant⁻¹ (−40.9 %; Fig. 3). All four doses of ZnO-NP significantly increased the fresh weight (9.106-9.612 g) and dry weight (1.068-1.160 g) without salt (Tukey 'a' across Z₁–Z₄). Under combined treatments, biomass recovery again peaked at S+Z₃, which attained 7.835 g fresh weight (90.6 % of C) and 0.953 g dry weight (89.1 % of C), significantly higher than S+Z₁ (6.031 g and 0.754 g) and only slightly above S+Z₂ (7.316 g and 0.849 g). The 100 mg L-1 dose was slightly lower than 75 mg L-1 for biomass, showing a similar height pattern (FW F-values 83.34, DW F-values 41.21, both P < 0.001).
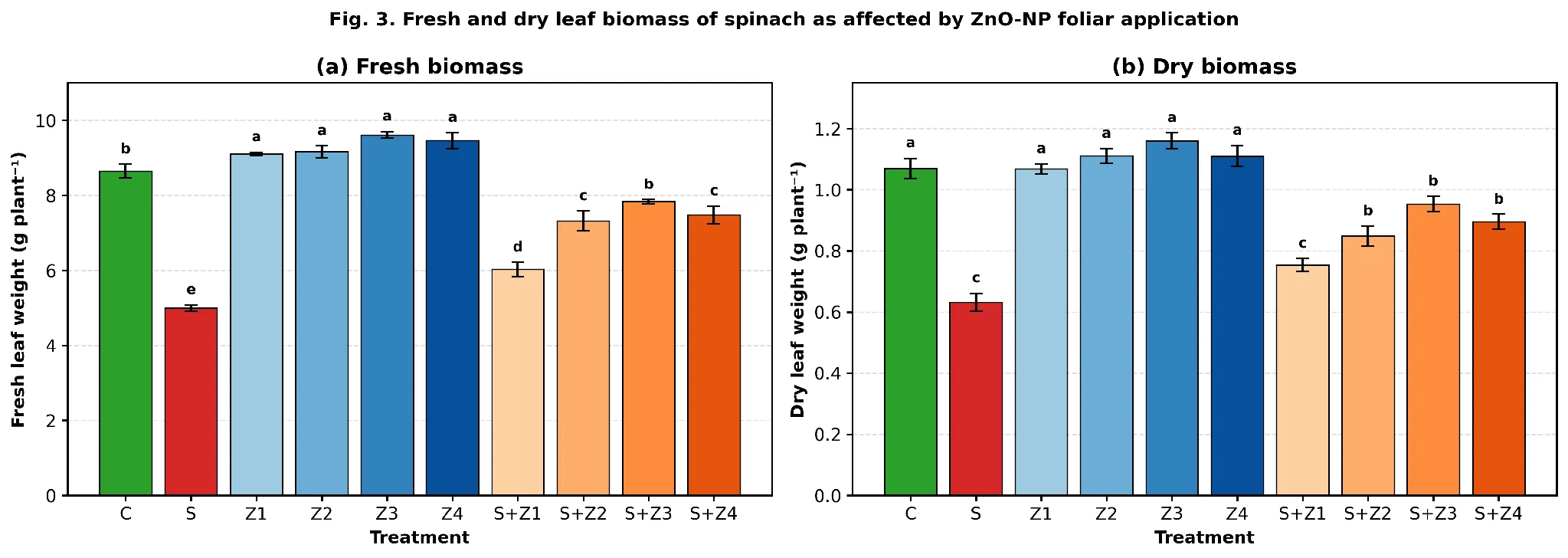
Fig. 3. (a) Fresh leaf weight and (b) dry leaf weight per plant of spinach as affected by foliar ZnO-NP application under control and 100 mM NaCl salt stress. Means ± SE (n = 3); different letters indicate significant differences (Tukey HSD, p < 0.05).
3.2 Chlorophyll content and photosystem II efficiency
Both the SPAD chlorophyll index and total chlorophyll content were significantly decreased when subjected to salt stress. SPAD declined from 45.8 in C to 30.2 in S (−34.1 %), while total chlorophyll dropped from 2.894 to 1.699 mg g⁻¹ FW (−41.3 %; Fig. 4). Under non-stressed conditions, the impact of ZnO-NPs gradually increased, reaching its highest value at 75 mg L⁻¹ (SPAD 51.4, total chlorophyll 3.327 mg g⁻¹ FW). Under saline conditions, S+Z₃ exhibited the highest recovery of SPAD (42.4 = 92.6 % of C; Tukey 'b') and total chlorophyll (2.641 mg g⁻¹ FW = 91.3 % of C). There was no clear relation between higher doses of Zn and better recovery in terms of SPAD (slightly better recovery for 100 mg L⁻¹ dose, F = 52.96, p < 0.001) and chlorophyll (marginally better recovery for 100 mg L⁻¹ dose, F = 55.84, p < 0.001).
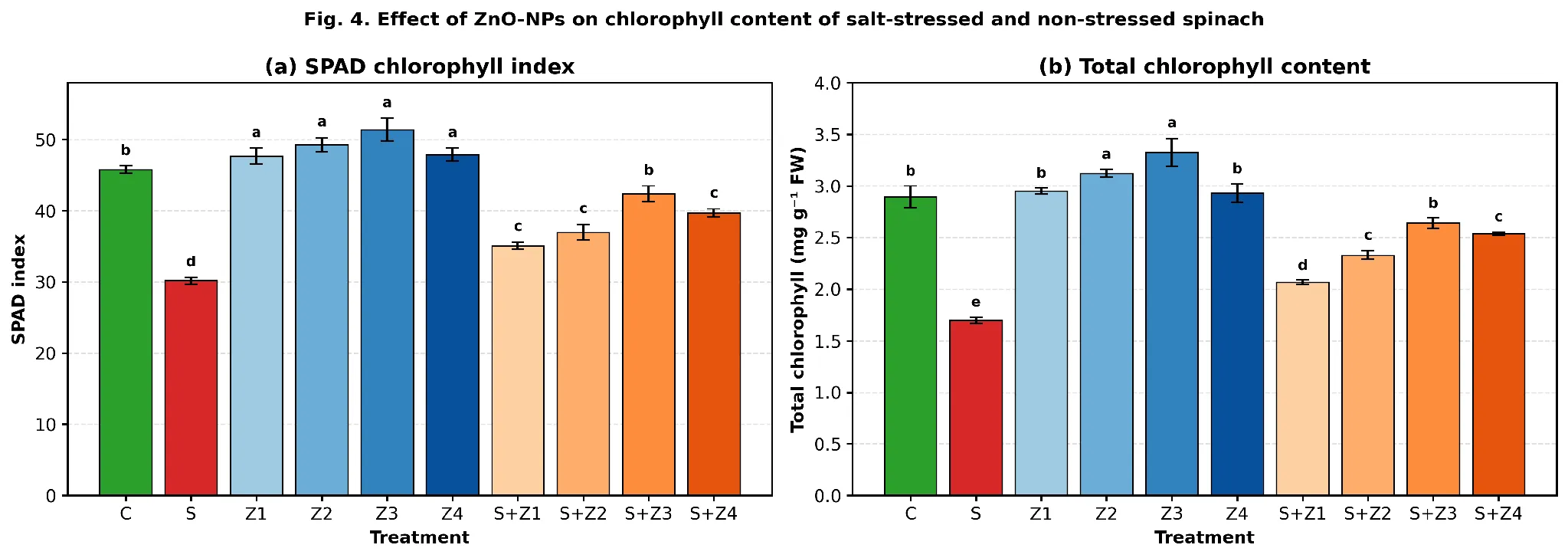
Fig. 4. Effect of foliar ZnO-NPs on (a) SPAD chlorophyll index and (b) total chlorophyll content (mg g⁻¹ FW) of spinach under control and salt-stressed conditions. Bars represent means ± SE (n = 3); different letters indicate significant differences at p < 0.05.
The PS II maximum quantum yield (Fv/Fm) was reduced from 0.827 (C) to 0.658 (S), which is a clear sign of photoinhibition (Fig. 5). Under non-stressed conditions all doses of ZnO-NP restored Fv/Fm to values of 0.840–0.863 (Z₁–Z₄, Tukey 'a'). Fv/Fm rose with increasing salinity from 0.715 (S+Z₁) to 0.784 (S+Z₃) which is approximately 94.8 % of the control, and 0.780 at the highest salinity (S+Z₄). The overall model was highly significant (F = 143.51; p = 3.54 × 10⁻¹⁶).
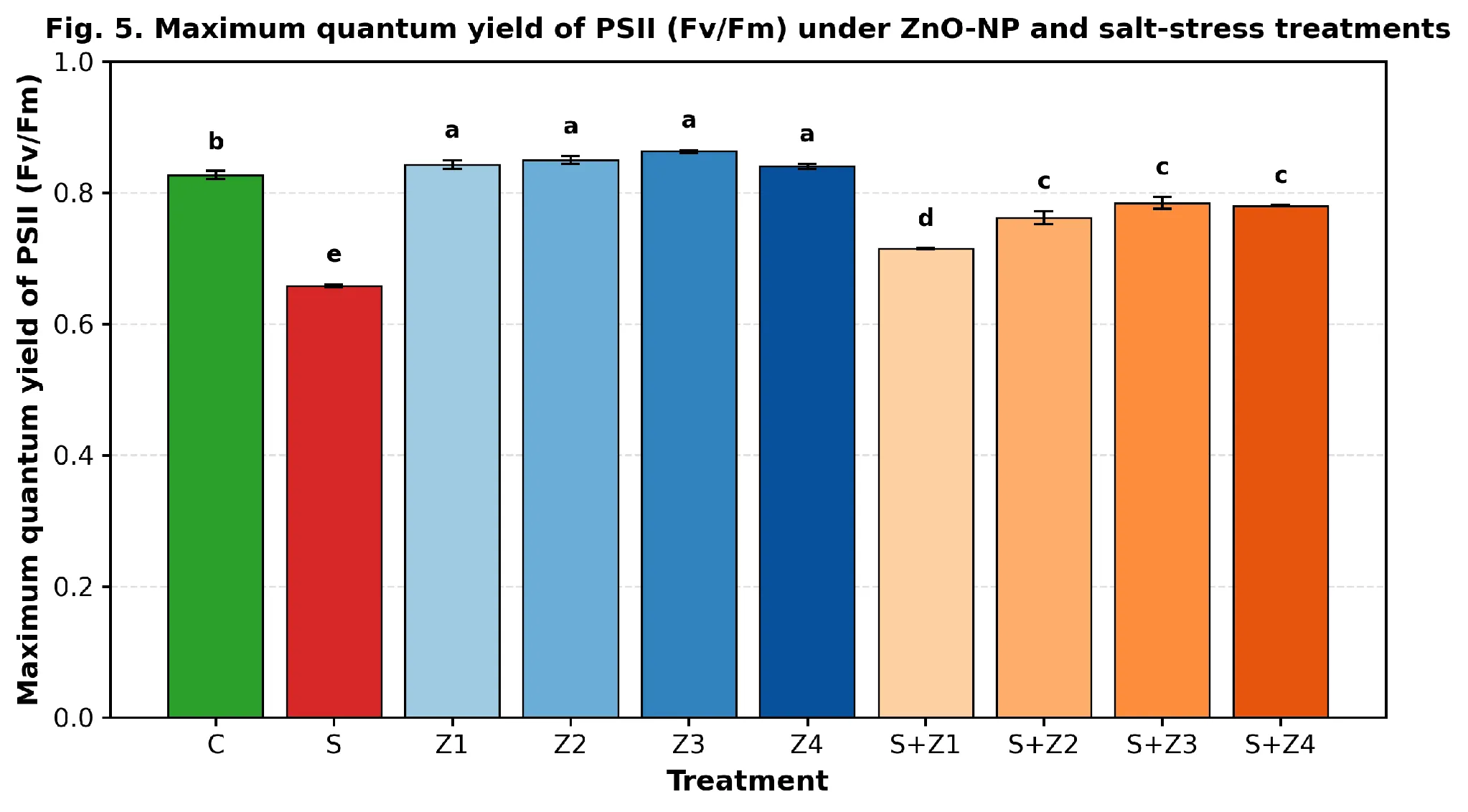
Fig. 5. Maximum quantum yield of photosystem II (Fv/Fm) in spinach leaves treated with foliar ZnO-NPs under control and salt-stressed conditions. Means ± SE (n = 3); different letters indicate significant differences (Tukey HSD, p < 0.05).
3.3 Water relations and membrane integrity
When plants were under severe osmotic stress (S), relative water content dropped significantly from 87.5 % in C to 61.7 % (Fig. 6a). The application of ZnO-NPs further enhanced RWC value by 88.2–91.1 % in the non-stressed treatments. With salinity, RWC dose-dependently increased from 70.1 % (S+Z₁) to maximum 82.2 % (S+Z₃; ≈94 % of the control), and 78.8 % was obtained with S+Z₄. The rate of electrolyte leakage, which is a measure of the integrity of the membrane, increased sharply from 12.35 % in C to 42.43 % in S (Fig. 6b). The EL was slightly reduced by foliar ZnO-NPs alone (11.36 to 11.82 %). Under combined treatments, EL declined progressively to a minimum of 18.53 % at S+Z₃ — a 56.3 % reduction relative to S — and was 22.33 % at S+Z₄ (F = 122.4 for RWC; F = 254.8 for EL; both p < 0.001).
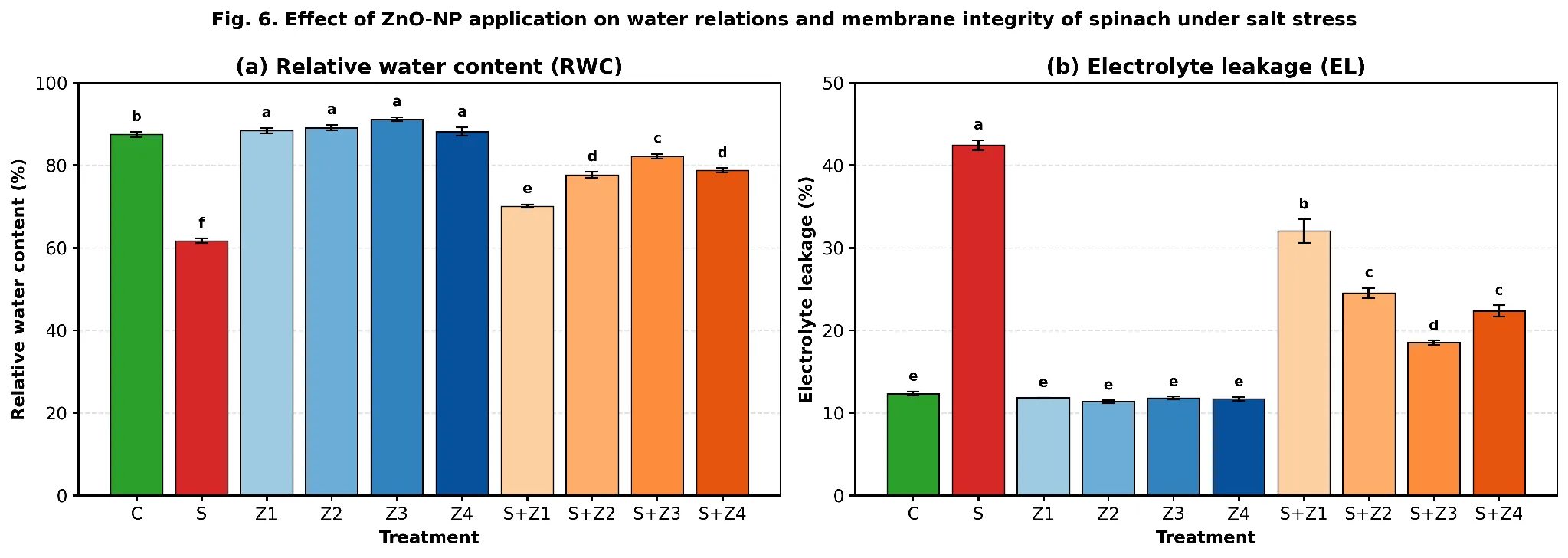
Fig. 6. (a) Relative water content (RWC, %) and (b) electrolyte leakage (EL, %) of spinach leaves in response to foliar ZnO-NPs under control and 100 mM NaCl salt stress. Means ± SE (n = 3); different letters indicate significant differences (Tukey HSD, p < 0.05).
3.4 Ionic homeostasis: Na⁺, K⁺ and K⁺/Na⁺ ratio in shoots
Salinity sharply increased shoot Na⁺ from 2.12 mg g⁻¹ DW in C to 19.51 mg g⁻¹ DW in S (a 9.2-fold increase) and reduced shoot K⁺ from 33.54 to 17.78 mg g⁻¹ DW (Fig. 7). In the shoot, the K⁺/Na⁺ ratio that serves as an important marker of salt tolerance dropped from 15.82 in C to 0.91 in S (Table 1; Fig. 7c). This imbalance was significantly decreased by ZnO-NPs applied to the leaves. Without the presence of salt, there was a slight increase in K⁺ accumulation by ZnO-NPs (34.78–36.49 mg g⁻¹ DW; Tukey 'a') but no changes to Na⁺. Increasing the dose of ZnO-NP under salinity progressively lowered Na⁺ accumulation from 14.79 (S+Z₁) to 8.09 mg g⁻¹ DW (S+Z₃) which is a 58.5 % reduction compared to S, and increased K⁺ from 24.61 to 31.34 mg g⁻¹ DW (S+Z₃) (76.3 % recovery toward C). Consequently, the K⁺/Na⁺ ratio improved from 1.66 (S+Z₁) to 3.87 (S+Z₃), a 4.25-fold increase over S. The dose of 100 mg L-1 yielded intermediate results (Na+ = 9.60 mg g-1 DW, K+ = 30.19 mg g-1 DW and K+/Na+ = 3.15), which demonstrates that the dose of 75 mg L-1 is the most beneficial for ionic-homeostasis.
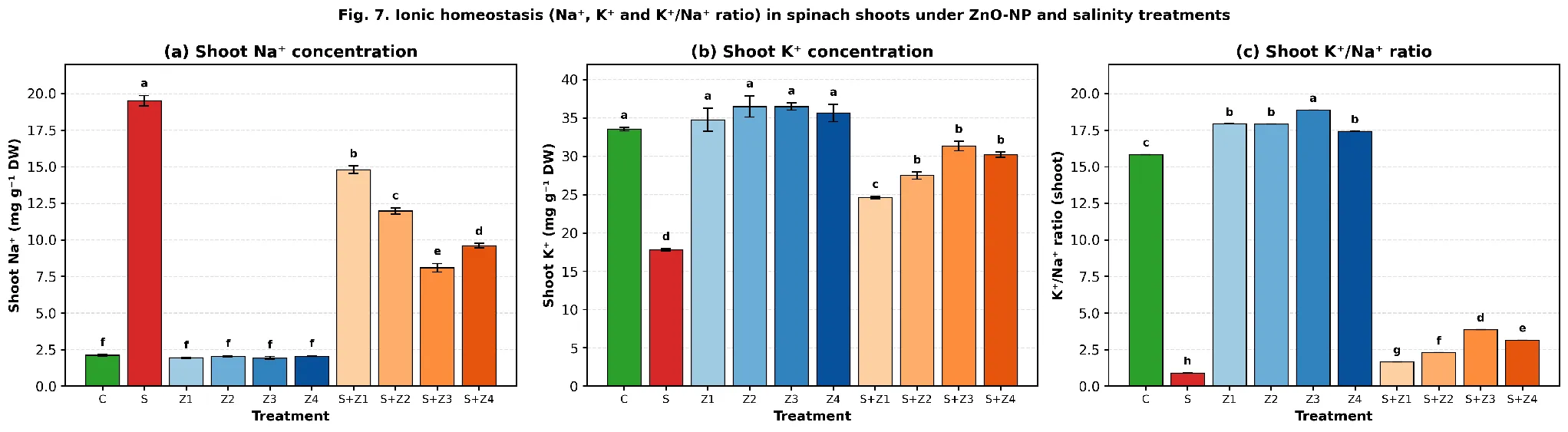
Fig. 7. Shoot (a) Na⁺ concentration, (b) K⁺ concentration and (c) K⁺/Na⁺ ratio of spinach in response to foliar ZnO-NP application under control and salt-stress conditions. Means ± SE (n = 3); different letters indicate significant differences (Tukey HSD, p < 0.05).
Table 1. Mean ± SE of all measured growth, physiological and ionic parameters across treatments (n = 3).
|
Parameter |
C |
S |
Z₁ |
Z₂ |
Z₃ |
Z₄ |
S+Z₁ |
S+Z₂ |
S+Z₃ |
S+Z₄ |
|
Plant height (cm) |
28.33 ± 0.24 |
17.86 ± 0.36 |
30.05 ± 0.61 |
30.66 ± 0.36 |
30.39 ± 0.76 |
29.25 ± 0.41 |
20.51 ± 0.64 |
22.81 ± 0.37 |
25.01 ± 0.33 |
23.93 ± 0.25 |
|
Leaf area (cm²) |
93.71 ± 3.06 |
52.20 ± 1.58 |
91.26 ± 2.77 |
101.94 ± 0.83 |
101.90 ± 2.01 |
98.26 ± 2.45 |
64.25 ± 2.05 |
74.02 ± 1.31 |
84.19 ± 2.26 |
79.37 ± 1.30 |
|
Fresh leaf wt (g) |
8.647 ± 0.187 |
4.991 ± 0.086 |
9.106 ± 0.037 |
9.166 ± 0.156 |
9.612 ± 0.084 |
9.453 ± 0.220 |
6.031 ± 0.189 |
7.316 ± 0.259 |
7.835 ± 0.051 |
7.476 ± 0.238 |
|
Dry leaf wt (g) |
1.070 ± 0.033 |
0.632 ± 0.029 |
1.068 ± 0.016 |
1.111 ± 0.024 |
1.160 ± 0.026 |
1.110 ± 0.035 |
0.754 ± 0.021 |
0.849 ± 0.033 |
0.953 ± 0.026 |
0.896 ± 0.024 |
|
SPAD index |
45.8 ± 0.5 |
30.2 ± 0.5 |
47.7 ± 1.1 |
49.3 ± 1.0 |
51.4 ± 1.6 |
47.9 ± 0.9 |
35.1 ± 0.5 |
37.0 ± 1.1 |
42.4 ± 1.1 |
39.7 ± 0.6 |
|
Total chlorophyll (mg g⁻¹ FW) |
2.894 ± 0.105 |
1.699 ± 0.029 |
2.951 ± 0.029 |
3.123 ± 0.038 |
3.327 ± 0.135 |
2.932 ± 0.090 |
2.065 ± 0.022 |
2.330 ± 0.043 |
2.641 ± 0.049 |
2.537 ± 0.015 |
|
Fv/Fm |
0.827 ± 0.006 |
0.658 ± 0.002 |
0.843 ± 0.007 |
0.850 ± 0.006 |
0.863 ± 0.002 |
0.840 ± 0.004 |
0.715 ± 0.001 |
0.762 ± 0.010 |
0.784 ± 0.009 |
0.780 ± 0.001 |
|
RWC (%) |
87.5 ± 0.6 |
61.7 ± 0.6 |
88.4 ± 0.6 |
89.1 ± 0.6 |
91.1 ± 0.5 |
88.2 ± 1.0 |
70.1 ± 0.4 |
77.7 ± 0.7 |
82.2 ± 0.5 |
78.8 ± 0.5 |
|
Electrolyte leakage (%) |
12.35 ± 0.26 |
42.43 ± 0.62 |
11.82 ± 0.03 |
11.36 ± 0.19 |
11.80 ± 0.17 |
11.69 ± 0.25 |
32.06 ± 1.44 |
24.54 ± 0.61 |
18.53 ± 0.29 |
22.33 ± 0.69 |
|
Shoot Na⁺ (mg g⁻¹ DW) |
2.12 ± 0.06 |
19.51 ± 0.35 |
1.94 ± 0.03 |
2.04 ± 0.04 |
1.93 ± 0.09 |
2.05 ± 0.02 |
14.79 ± 0.26 |
11.97 ± 0.21 |
8.09 ± 0.29 |
9.60 ± 0.16 |
|
Shoot K⁺ (mg g⁻¹ DW) |
33.54 ± 0.21 |
17.78 ± 0.16 |
34.78 ± 1.49 |
36.49 ± 1.38 |
36.47 ± 0.46 |
35.64 ± 1.13 |
24.61 ± 0.13 |
27.50 ± 0.46 |
31.34 ± 0.62 |
30.19 ± 0.34 |
|
K⁺/Na⁺ ratio |
15.82 |
0.91 |
17.93 |
17.92 |
18.87 |
17.42 |
1.66 |
2.30 |
3.87 |
3.15 |
Note: Values are means ± SE of three independent replicates. Treatment codes: C = non-stressed control; S = 100 mM NaCl; Z₁–Z₄ = 25, 50, 75 and 100 mg L⁻¹ ZnO-NPs (no salt); S+Z₁–S+Z₄ = corresponding ZnO-NP doses combined with 100 mM NaCl. All parameters showed treatment effects significant at p < 0.001 (two-way ANOVA).
4. DISCUSSION
From present study it is evident that foliar application of ZnO-NPs prepared using neem extract shows alleviating effect on spinach under several stresses such as 100 mM NaCl and 75 mg L⁻¹ (S+Z₃) is the optimal protective dose. The coordinated changes in growth, photosynthesis, water relations, membrane integrity and ionic balance suggest that the protective effect is mechanistically complex, involving more than one pathway. Spinach plants were shorter, had smaller leaves, less fresh and dry weight, as compared to the control, and this was consistent with the reported reduction in yield of leafy vegetables ranging from 30–50 % for exposure to 80–120 mM NaCl (Ors & Suarez, 2017; Saleem et al., 2021). This growth suppression can be explained by osmotic-phase reductions in turgor-driven cell expansion, decreased photosynthesis and by Na⁺-induced cytotoxicity which diverts carbon and energy from growth to stress defence (Munns & Tester, 2008). The treatment with ZnO-NP over the time gradually ameliorated these effects and the value of plant height, leaf area, fresh weight and dry weight was restored to 88 %, 90 %, 91 % and 89 % of the non-stressed, respectively. The agronomic significance of this recovery is significant given that the economic returns of a 10–20 % recovery in salt-affected horticulture can mean the difference between profitable and unprofitable seasons.
This growth response is the centerpiece of Zn nutrition. Zn²⁺ serves as a structural and catalytic cofactor of over 300 enzymes including: carbonic anhydrase, Cu/Zn superoxide dismutase, alcohol dehydrogenase and RNA polymerase, and is needed for biosynthesis of tryptophan, the precursor of the plant growth hormone indole-3-acetic acid (IAA) (Cakmak, 2000; Sturikova et al., 2018). Zn nutrition in this case thus stimulates cell elongation, leaf expansion and meristematic activity. The biphasic dose–response nature of ZnO-NP, with the lowest dose (25 mg L⁻¹) having lower efficacy and the highest dose (100 mg L⁻¹) showing slightly lower protection compared to the medium dose (75 mg L⁻¹), suggests that ZnO-NP action could be beneficial at low–medium doses and that higher doses might exhibit mild phytotoxicity due to the production of ROS or excess accumulation of Zn²⁺ as reported by Du et al. (2019) and Singh et al. (2022). In the case of S+Z₃, the SPAD index, total chlorophyll and Fv/Fm showed a strong recovery, highlighting the ability of ZnO-NPs to preserve the integrity of PSII reaction centre and the chlorophyll pigment pool. The loss of chlorophyll is caused by activation of chlorophyllase, replacement of Mg²⁺ in the porphyrin ring and ROS-mediated peroxidation of thylakoid lipids (Ashraf & Harris, 2013; Hasanuzzaman et al., 2020). The microelement Zn²⁺ provided by foliar ZnO-NPs helped stabilize the structure of the thylakoid membrane, increase the amount of chlorophyll in plants (increase in the synthesis of δ-aminolevulinic acid), and protect PSII from the effect of photoinhibition by activating Cu/Zn-SOD in the chloroplast (Cakmak, 2000; Sturikova et al., 2018; Rizwan et al., 2019). The recovery of Fv/Fm to 0.784 with S+Z₃ (close to the healthy plant value of ~0.83) is highly indicative of the preservation of the donor and acceptor side of PSII.
This clear improvement in RWC (from 61.7 % in S to 82.2 % in S+ZnO-NPs) along with the corresponding reduction in EL (from 42.4 % to 18.5 % in S+ZnO-NPs) shows the role/benefits of ZnO-NPs in maintaining water status of the cells and their membrane integrity under salinity. There are 2 possible mechanisms. Firstly, ZnO-NPs increase the leaf water potential through accumulation of compatible solutes like proline, glycine betaine and soluble sugars, and thus, enhance the turgor (Faizan et al., 2021). Second, Zn has been reported to stabilize biomembranes in terms of both structure and function by binding to the sulfhydryl groups of phospholipids and by activating Cu/Zn-SOD which decreases the lipid peroxidation of biomembranes (Cakmak, 2000; Alharby et al., 2016). This dual stabilizing effect is reflected in the strong inverse relationship between EL and RWC across treatments. The most remarkable effect of ZnO-NPs was on ionic homeostasis. Na⁺ content increased ~9 fold and K⁺ content reduced by ~47 % in shoots, resulting in a decrease in the K⁺/Na⁺ ratio from 15.82 to 0.91. It is widely accepted that the maintenance of a high cytosolic K+/Na+ ratio is the most important physiological determinant of salt tolerance and that cytosolic Na+ is required for stomatal regulation and for protein synthesis while excessive cytosolic Na+ interferes with these processes by competing with K+ for binding sites (Munns & Tester, 2008; Shabala & Pottosin, 2014). The foliar application of ZnO-NPs gradually reduced shoot Na⁺ (by 24, 39, 59 and 51 % at S+Z₁, S+Z₂, S+Z₃ and S+Z₄, respectively) while simultaneously increasing shoot K⁺ which led to a more than 4-fold increase in K⁺/Na⁺ ratio at 75 mg L⁻¹. The mechanism is likely due to the interaction of Zn with the plasma membrane H⁺-ATPase and Na⁺/H⁺ antiporters (SOS1 family) that energize Na⁺ extrusion and vacuolar sequestration, and protect Na+-uptake channels (HAK/AKT1) from damage by Na⁺ (Shabala & Pottosin, 2014; Faizan et al., 2021).
These results are put in perspective with the literature. Various crop species such as wheat (Alharby et al., 2016), tomato (Faizan et al., 2021), maize (Zafar et al., 2016), rice (Rizwan et al., 2019) and mung bean (Venkatachalam et al., 2017) were used in these studies, and the effective doses of foliar applications were generally in the range of 50–100 mg L⁻¹. The results in this study confirmed that the 75 mg L-1 dose was the optimal dose for spinach, in line with these reports, and strengthened the growing consensus that intermediate doses of ZnO-NPs are most effective, whereas higher doses can cause nano-specific phytotoxicity (Du et al., 2019; Singh et al., 2022). One of the key strengths of this study is the use of the green synthesized ZnO-NPs using an aqueous extract of Azadirachta indica (neem). Neem extract contains a combination of Zn²⁺ reductants and natural capping/stabilizing agents, which are nimbin, azadirachtin, polyphenols, flavonoids and terpenoids (Elumalai et al., 2015; Bhuyan et al., 2015). It avoids the use of potentially toxic chemical reductants (NaBH₄, hydrazine), low energy consumption (no high temperature calcination) and results in particles with biologically attached biomolecules that are more biocompatible with plant tissues than chemically synthesized NPs (Iravani, 2011; Singh et al., 2018; Madan et al., 2016). The capping biomolecules can also impart other biological activities such as antioxidants and anti-fungals; neem-derived phenolics are such also, and may exert synergistic activity as seen here.
Combined, the results point towards an integrated effect of foliar ZnO-NPs, where (i) the provision of bioavailable Zn for enzyme activation and chlorophyll biosynthesis, (ii) the stabilization of biomembranes and PSII from oxidative damage, (iii) the enhancement of osmotic adjustment and water relations, and (iv) the restoration of K⁺/Na⁺ homeostasis by activation of the ion-transport machinery. The biphasic dose response further confirmed that the dose range 25–75 mg L⁻¹ was sub-optimal, and 100–125 mg L⁻¹ was approaching the toxicity limit. The consequences are very real. Worldwide, around 20 – 33 % of irrigated agricultural land is saline (Hassani et al., 2021) and smallholder producers of leafy vegetables are particularly impacted by salinity in Asia and Africa. The technology is accessible, low cost and sustainable as it requires only a few hundred millilitres of suspension per hectare of foliar sprays of ZnO-NP at the rate of 75 mg L⁻¹ which can be prepared locally using neem leaves. The successful adoption of this technology will require field testing over a range of cultivars, soil and salinity conditions as well as assessing the trophic transfer and consumer safety of Zn in edible leaves. While these are positive outcomes, there are some potential areas for further study and limitations that should be noted. The experiment was carried out in controlled greenhouse environment for one growing cycle and field studies are recommended to verify the efficacy of the ZnO-NPs under various environmental conditions and extended crop cycles. The action of ZnO-NPs on the expression of salt-stress responsive genes (SOS1, HKT1, NHX1) and transcripts of enzymes involved in the scavenging of ROS should be investigated further to understand the molecular mechanism of salt tolerance. Finally, prior to commercial recommendation, extensive risk assessment of the translocation of Zn to edible leaves must be undertaken, including consumer exposure modelling.
CONCLUSION
This study shows that foliar application of zinc oxide nanoparticles synthesized using neem extract is an excellent environmentally-friendly method for mitigating the adverse effects of salinity (100mM NaCl) on spinach (Spinacia oleracea L.). Under salt stress, the height, leaf area, fresh biomass, dry biomass, SPAD index, total chlorophyll, electrolyte leakage, and RWC were reduced by 37, 44, 42, 41, 34, 41, 3.4 and 9.2-fold, respectively, while shoot Na⁺ increased by 3.4-fold and shoot K⁺/Na⁺ ratio decreased from 15.82 to 0.91.
All of these effects were strongly mitigated in a dose-dependent fashion by foliar sprays of ZnO-NP. The dose of 75 mg L⁻¹ (S+Z₃) was the most effective and normalized about 88 % of the plant height, 90 % of the leaf area, 91 % of fresh biomass, 89 % of dry biomass, the SPAD index to 92.6 %, total chlorophyll to 91.3 %, the Fv/Fm index to 0.784, and the RWC to 82.2 % of the non-stressed control among the four tested doses. This same dose reduced electrolyte leakage by 18.5 % and decreased shoot Na⁺ by about 59% while increasing shoot K⁺ by about 76 % and increasing the shoot K⁺/Na⁺ ratio over four-fold, bringing it closer to that of healthy plants. The 100 mg L-1 showed slightly less protection than 75 mg L-1 suggesting a biphasic response and saturation/mild-toxicity threshold at > 75 mg L-1.
Based on the agronomic point of view, it was concluded that 75 mg L⁻¹ was the recommended foliar application rates of ZnO-NPs in spinach under saline conditions. The synthesis of nanoparticles using the neem leaf extract is an inherently sustainable, environmentally friendly and locally feasible process as opposed to synthetic nanomaterials, which is in tune with the green chemistry and circular bio-economy. Preparation of the nanoparticles using neem phytochemicals as Zn²⁺ reductants as well as biocompatible capping agents will very likely improve the colloidal stability and biological activity of the prepared nanoparticles.
The results of this study should be further confirmed by multi-location field trials on different spinach cultivars and salinity levels, dose response studies under combined biotic and abiotic stresses, molecular and transcriptomic analysis of plants' response to foliar application of ZnO-NP and thorough assessment of Zn translocation, residues and consumer safety in edible leaves. Co-application of other complementary technologies such as biofertilizers, plant growth promoting rhizobacteria or other green-synthesized nano-formulation may further boost the resilience of leafy vegetable production on the increasing salt-affected lands of the world.
Acknowledgement: Authors would like to acknowledge Jayoti Vidyapeeth Women's University for providing lab access and resources.
Conflict of Interest
The authors declare no competing financial interests or conflicts of interest related to this work.
REFERENCES

Jyoti Rani, Neeraj Choudhary, Foliar Application of Neem-Mediated Zinc Oxide Nanoparticles Alleviates Salinity Stress in Spinach (Spinacia oleracea L.): Improvements in Growth, Photosynthetic Efficiency, Water Relations, and Ionic Homeostasis, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 7, 688-703, https://doi.org/10.5281/zenodo.21156875
 10.5281/zenodo.21156875
10.5281/zenodo.21156875