
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
1Research Scholar, School of Pharmacy, P Savani University, Kosamba, Suraj, Gujrat, India-394125
2Research Scholar, School of Pharmacy, P Savani University, Kosamba, Suraj, Gujrat, India-394125
3Professor, Department of Pharmaceutics, School of Pharmacy, P P Savani University, Kosamba, Suraj, Gujrat, India-394125
4 Professor and Principal, Department of Pharmaceutics, School of Pharmacy, P P Savani University, Kosamba, Suraj, Gujrat, India-394125
Dry eye disease (DED), clinically designated keratoconjunctivitis sicca, is a debilitating multifactorial disorder of the ocular surface that afflicts an estimated 1.5 billion individuals globally. Its pathogenesis is fundamentally rooted in a self-perpetuating metabolic and oxidative crisis within corneal epithelial cells, wherein chronic inflammation progressively depletes intracellular nicotinamide adenine dinucleotide phosphate (NADPH) and adenosine triphosphate (ATP), dismantling the cell's antioxidant and regenerative capacity. Current pharmacological interventions, including cyclosporine A (Restasis) and lifitegrast (Xiidra), target downstream immune effectors but do not directly rectify the bioenergetic deficit that underlies corneal cell deterioration. In this context, an entirely unprecedented cross-kingdom bioengineering strategy has emerged from the National University of Singapore, offering a conceptually transformative approach to DED therapy. Researchers isolated structurally intact thylakoid grana nanoparticles from spinach (Spinacia oleracea) leaves using a patented mild mechanical and chemical extraction protocol, yielding sub-micron photosynthetic particles designated LEAF (Light-reaction Enriched thylAkoid NADPH-Foundry). These nano-thylakoids, approximately 400 nanometres in diameter, are specifically engineered to retain the complete thylakoid electron transport chain while eliminating the Calvin-cycle stroma that would otherwise consume the very NADPH being generated. Once administered as eye drops and internalised by corneal cells, LEAF particles function as plug-and-play neo-organelles, autonomously harvesting ambient visible light to drive photosynthetic electron flow, regenerating NADPH at rates approximately twenty percent greater than unencapsulated thylakoids and restoring ATP. In preclinical mouse models, LEAF eye drops reversed corneal damage to near-healthy tissue levels within five days, outperforming cyclosporine A. Ex vivo experiments demonstrated greater than ninety-five percent reduction in hydrogen peroxide in patient tear samples and a twenty-fold increase in NADPH levels. This review comprehensively analyses the molecular pathophysiology underpinning LEAF's therapeutic rationale, the photosynthetic mechanisms responsible for its efficacy, its formulation science, proposed optimisation strategies beyond existing publications, critical safety considerations, and the regulatory and translational pathway toward clinical application.
The ocular surface is one of the most active and immunologically distinct areas in man. It must remain optically clear and maintain the cellular homeostasis despite environmental challenges like exposure to ultraviolet radiation, air borne particulates, antigens from microbes and osmotic stresses due to evaporation, with remarkable precision. [1,2] It is also one of the most common conditions that impact this highly complex system, known as dry eye disease (DED). DED is defined as a multifactorial disease of the ocular surface caused by the loss of tear film homeostasis, which includes tear film hyperosmolarity, tear film instability, neurogenic inflammation and epithelial damage, resulting from a complex interplay of these factors. [3, 4] DED is devastating, with a worldwide impact. More than 1.5 billion people around the world are estimated to be affected, and the prevalence of the disease could be as high as 50% or as low as 5% depending on the diagnostic criteria, geographic region and population age.[5,6] DED prevalence in India has been reported from 18.4% to 32.5% in urban population and the various fastest increasing risk factors are digital device overuse, environmental pollution and wearing contact lenses (CLs).[7,8] The direct and indirect estimated annual cost burden from DED in the USA is just over USD 55 billion per year, which includes the cost of prescription medicines, loss of productivity and impairment to vision related quality of life. [9,10] Clinical research over the last few years has not significantly advanced the therapeutic armamentarium for DED, which is still largely symptomatic. Eye drops that help lubricate the surface of the eye do only a quick fix for surface lubrication.[11] Cyclosporine A (0.05%, Restasis, Allergan) is an antibody directed against the T lymphocytes and when applied to the eyes, takes three to six months to have any clinical effect, but has been found to cause significant discomfort on instillation. [12,13] Again, Lifitegrast (5%, Xiidra, Novartis) is a competitive inhibitor of the binding of the lymphocyte function-associated antigen-1 (LFA-1) to intercellular adhesion molecule-1 (ICAM-1); thus, it dampens the inflammatory response of the eyes by inhibiting T-cell activation.[14,15] Both agents fail to replenish the intracellular pools of both NADPH and ATP that are important in the progressive loss of an antioxidant defence of corneal epithelial cells.[16,17] A self-reinforcing metabolic exhaustion and oxidative stress loop is central to the pathogenic molecular basis of DED. The oxidative stress caused by chronic inflammation at the ocular surface results in the formation of reactive oxygen species (ROS) which are overwhelming endogenous antioxidants, mainly NADPH-dependent glutathione reductase and thioredoxin reductase pathway.[18,19] The corneal cell will lose its ability to scavenge ROS as a consequence of the increased consumption of NADPH by the pentose phosphate pathway, thus leading to further inflammation, goblet cell loss and cellular apoptosis.[20,21] This vicious cycle is exacerbated by depletion of ATP, which is required for maintaining ionic gradients as well as for repair of membranes and secretion of mucus.[22,23] There is no current approved therapy that is directly aimed at this bioenergetic collapse.[24,25] In this context, the development of the concept of transfer of photosynthesis system from plants to mammals is a conceptual revolution of the highest order. A team of researchers led by Associate Professor David Leong Tai Wei and the first author of the study Dr Xing Kuoran of the National University of Singapore (NUS) have managed to successfully engineer LEAF (Light-reaction Enriched thylAkoid NADPH-Foundry) nanoparticles based on spinach (Spinacia oleracea) thylakoid grana.[26,27] This strategy has been shown to be effective in a preclinical model, better than cyclosporine A, and to restore corneal integrity to close to normal within five days, after incorporation in the cells of the cornea.[28] This review aims to provide a thorough scholarly examination of the theoretical and experimental basis for LEAF technology, its formulation science, mechanistic dual-pathway action, proposed novel optimisation strategies, safety profile, regulatory outlook, and the broader implications of cross-kingdom organelle transplantation as a therapeutic principle. The discussion includes several original proposals not documented in the existing literature, including surface-functionalized mucoadhesive LEAF variants, photo-enhancing ocular insert systems, Nrf2 co-activator co-encapsulation, and cryo-preserved reconstitutable formulations, designed to maximise the clinical translatability of this technology.
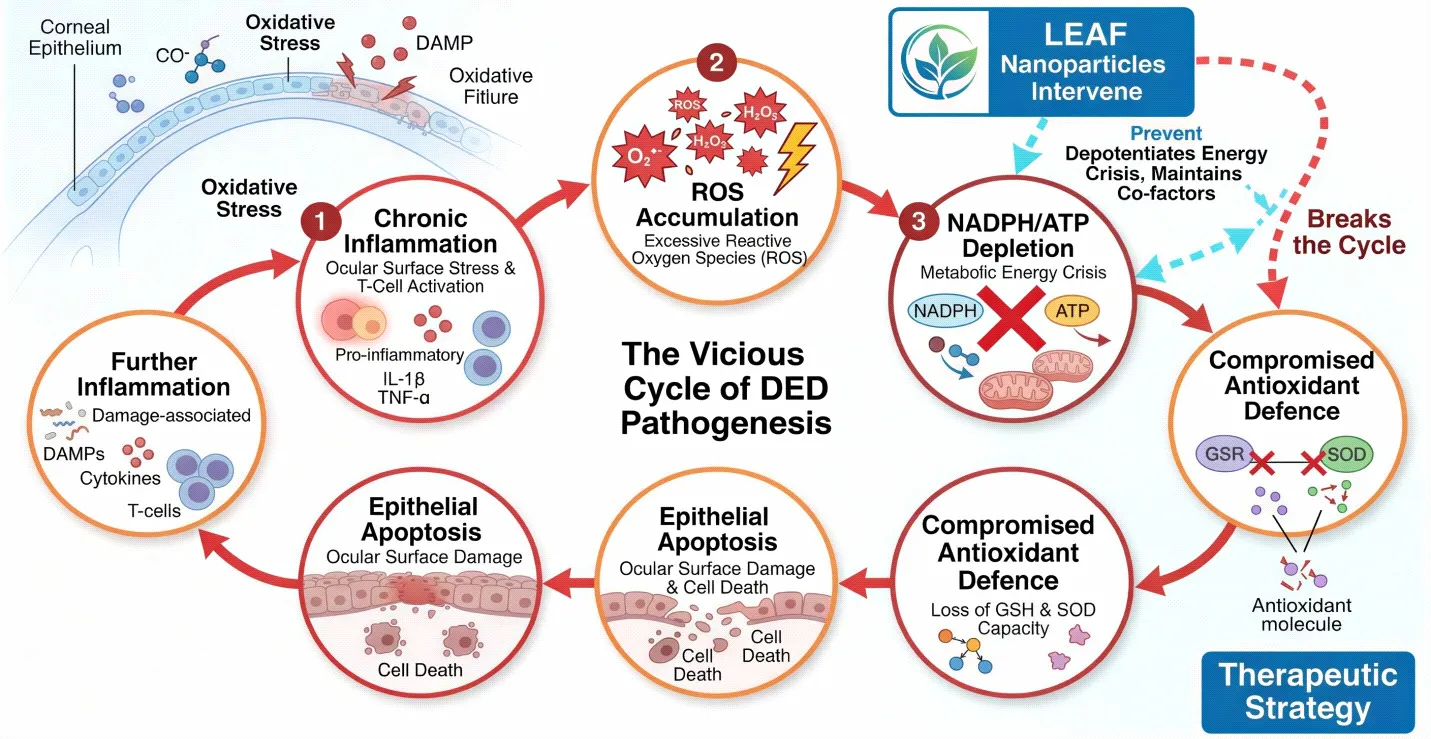
Fig. 1: Schematic representation of the vicious cycle of DED pathogenesis: Chronic inflammation → ROS accumulation → NADPH/ATP depletion → compromised antioxidant defence → epithelial apoptosis → further inflammation. LEAF nanoparticles intervene at the NADPH/ATP depletion node.
DED is classified into distinct phenotypic subtypes based on the predominant pathophysiological mechanism, as summarized below.
Table 1: Classification of Dry Eye Disease — Subtypes, Mechanisms, and Biomarkers
|
Type |
Mechanism |
Biomarkers |
Clinical Features |
|
Aqueous-deficient (ADDE) |
Lacrimal gland hyposecretion; reduced basal tear volume |
Low lactoferrin; elevated IL-6, IL-8 |
Schirmer test < 5 mm/5 min; low TBUT; keratitis sicca |
|
Evaporative (EDE) |
Meibomian gland dysfunction; lipid layer thinning; osmotic stress |
Elevated MMP-9; lipid layer < 60 nm; osmolarity > 316 mOsm/L |
Rapid TBUT (< 5 s); lid margin telangiectasia; chalazion |
|
Mixed (ADDE + EDE) |
Dual pathway; concurrent immune activation and lipid deficiency |
Elevated MMP-9, IL-1β, CXCL-10; low lactoferrin |
DEWS II grade 2–3; concurrent evaporative and secretory deficits |
|
Neuropathic ocular pain |
Peripheral/central sensitisation; altered TRPV1/TRPA1 signalling |
Abnormal IVCM morphology; elevated substance P |
Disproportionate pain; normal ocular surface signs; allodynia |
2. PATHOPHYSIOLOGY OF DRY EYE DISEASE: THE METABOLIC AND OXIDATIVE BASIS
2.1 Tear Film Architecture and Its Vulnerability
The human tear film is a trilaminar structure comprising an outermost lipid layer secreted by meibomian glands, an aqueous middle layer produced by the main and accessory lacrimal glands, and an innermost mucin-rich glycocalyx layer anchored by membrane-spanning mucins (MUC1, MUC4, MUC16) produced by conjunctival goblet cells.[29,30] Each layer serves distinct functions: the lipid layer retards evaporation, the aqueous layer carries oxygen, immunoglobulins, lactoferrin, and lysozyme to the avascular corneal epithelium, and the mucin layer reduces surface tension and promotes tear spreading over the hydrophobic glycocalyx.[31] The integrity of this structure is maintained through precise neurogenic reflex arcs and hormonal signalling, both of which are compromised in DED.[32,33]
In DED, disruption of any component initiates a cascade. This increases secretion via the activation of the mitogen-activated protein kinase (MAPK) pathway, nuclear factor kappaB (NF-κB) signalling and activation of the NLRP3 inflammasome in corneal epithelial cells and subsequent production of IL-1β,[34,35] IL-6, IL-8, TNF-α and MMP-9, further damaging the epithelial tight junction barrier, contributing to the hyperosmolarity.[36,37,38]
2.2 The NADPH-Centred Oxidative Crisis
The NADPH is located in the middle of the antioxidant network in the corneal epithelial cell. It also acts as the obligate reducing equivalent for the enzyme glutathione reductase to reduce oxidised glutathione (GSSG) to reduced glutathione (GSH), the most abundant intracellular peroxide-scavenging antioxidant, and is also a substrate for the different isoforms of NADPH oxidase (NOX) which are involved in regulated ROS production.[39,40] It is also the obligate reducing equivalent of the enzyme glutathione reductase which reduces oxidised glutathione (GSSG) to reduced glutathione (GSH), the most abundant intracellular peroxidase-scavenging antioxidant and a substrate for the various forms of NADPH oxidase (NOX), which are involved in controlled ROS formation.[41] In healthy corneal cells, NADPH is regenerated primarily through the pentose phosphate pathway (glucose-6-phosphate dehydrogenase reaction) and, to a lesser extent, by malic enzyme and isocitrate dehydrogenase.[42,43]
In DED, chronically elevated ROS generated by hyperosmolarity, UV exposure, and inflammatory cytokine signalling overwhelm the pentose phosphate pathway's regenerative capacity.[44,45] The resulting NADPH deficit creates a situation in which oxidised glutathione cannot be recycled, mitochondrial membrane potential collapses, cytochrome c is released, and apoptotic caspase cascades are activated.[46,47] Simultaneously, the depletion of ATP — normally generated by mitochondrial oxidative phosphorylation — disrupts Na⁺/K⁺-ATPase activity, cell volume regulation, and active transport of trefoil factor family peptides responsible for epithelial repair.[48,49] The corneal cell is thus locked in a state of bioenergetic insufficiency from which conventional anti-inflammatory therapy cannot rescue it because neither cyclosporine A nor lifitegrast replenishes these depleted cosubstrates.[50,51]
2.3 The Role of DUOX2, MMP-9, and Inflammatory Amplifiers
Recent mechanistic studies have identified dual oxidase-2 (DUOX2), a member of the NADPH oxidase family highly expressed in corneal and conjunctival epithelium, as a key amplifier of the oxidative cascade in DED.[52] TLR4-dependent DUOX2 activation generates superoxide and hydrogen peroxide in response to lipopolysaccharide and hyperosmolarity, promoting HMGB1 release, which in turn recruits macrophages and dendritic cells to the ocular surface.[53] Concurrently, MMP-9 (matrix metalloproteinase-9), whose expression is elevated in DED tear fluid, degrades tight-junction proteins (ZO-1, occludin) and E-cadherin, destabilising the epithelial barrier.[54,55] Cytokines including IL-17A (secreted by γδ T cells in the conjunctiva) further promote barrier disruption and reduce the expression of goblet-cell mucins.[56]
This multi-effector inflammatory milieu is not addressed by any single-target therapy. The LEAF approach, by replenishing NADPH inside the cell, simultaneously addresses the ROS crisis (by fuelling GSH recycling and ROS detoxification), restores ATP for active barrier repair, and exerts anti-inflammatory effects extracellularly through the capacity of released NADPH to scavenge hydrogen peroxide in the tear film.[57,58]
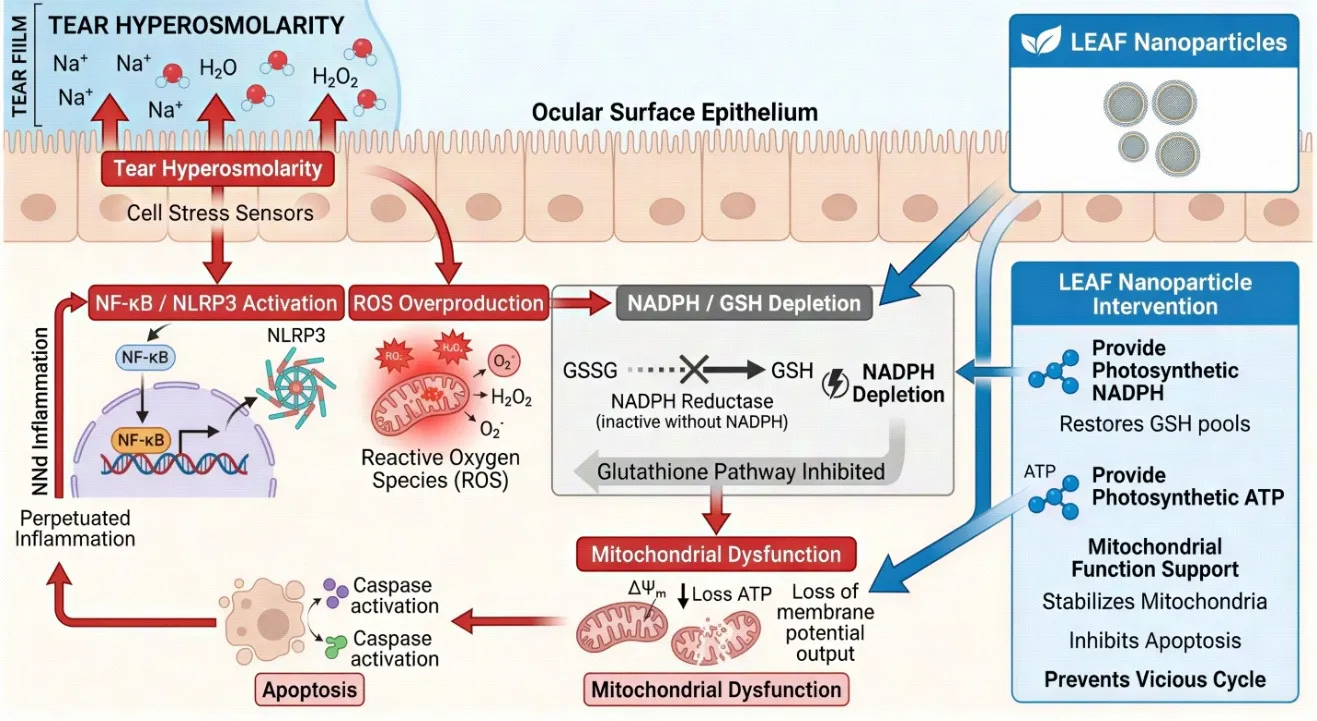
Fig. 2: Molecular pathophysiology of DED highlighting the NADPH depletion cycle: Tear hyperosmolarity → NF-κB / NLRP3 activation → ROS overproduction → NADPH/GSH depletion → mitochondrial dysfunction → apoptosis → perpetuated inflammation. LEAF nanoparticles intervene by providing photosynthetic NADPH and ATP.
3. PHOTOSYNTHETIC MACHINERY: THYLAKOID GRANA AS THE ACTIVE ENGINE
3.1 Structure and Function of Spinach Chloroplasts
Chloroplasts, the photosynthetic organelles of plant cells, are bounded by a double envelope membrane and contain an elaborate internal membrane system called the thylakoid network.[59,60] Within the thylakoid membrane are stacked disc-like structures called grana (singular: granum), which are the sites of the light-dependent reactions of photosynthesis.[61] Each granum stack contains multiple thylakoid discs (lamellae) connected by non-stacked stromal lamellae.[62] The grana membranes are densely packed with two principal photosynthetic protein supercomplexes: Photosystem II (PSII), which uses light energy to oxidise water (releasing O₂) and reduce plastoquinone, and Photosystem I (PSI), which reduces ferredoxin using electrons originating from PSII via the plastoquinone pool, the cytochrome b6f complex, and plastocyanin.[63,64]
The terminal step relevant to LEAF's therapeutic mechanism is the reaction catalysed by ferredoxin-NADP⁺ reductase (FNR): reduced ferredoxin (Fd_red) transfers electrons to NADP⁺, yielding NADPH.[65] This is the final photosynthetic electron acceptor in the non-cyclic (Z-scheme) electron transport chain. Simultaneously, the proton gradient (ΔpH) established across the thylakoid membrane by water oxidation and plastoquinol oxidation drives ATP synthase (the CF₀-CF₁ complex), generating ATP from ADP and inorganic phosphate.[66] Both products — NADPH and ATP — are normally consumed in the stroma by the Calvin–Benson–Bassham cycle for CO₂ fixation.[67,68]
The critical innovation of LEAF lies in the selective removal of the stroma (the site of Calvin cycle enzymes) while preserving the intact grana. Thus the thylakoid is a new particle for the production of NADPH and ATP only – there is no Calvin cycle to use up the products anymore – and the NUS study shows that the particle produces ~20% more NADPH than the unencapsulated crude thylakoid.[69,70]
3.2 Why Spinacia oleracea Outperforms Other Leafy Sources
The high yield of plant photosynthetic machinery per gram of leaf tissue, increase in the number of chloroplasts, increase in the number of chloroplast grana, and increase in PSII to PSI stoichiometry favouring non-cyclic electron transport pathways are all advantageous features of S. oleracea that were found in the systematic comparison and were also attributed to other advantages of S. oleracea, including its abundance in the market, familiarity as a food-grade plant, and well-characterised phytochemical profile.[71,72,73,74]
3.3 The Endosymbiotic and Cross-Kingdom Precedent
While a new idea for deliberately designed therapeutics, the idea of functional plant organelles within the animal cell finds a remarkable precedent in nature. Sustained kleptoplasty is known to occur in the sea-slugs of the genus Elysia, particularly Elysia chlorotica, and the chloroplasts (kleptoplasts) remain photosynthetically active for weeks to months, contributing to the slug's energy metabolism during starvation experiments. The genetic mechanism of sustained kleptoplasty is still under debate, while the biological principle (the ability of thylakoid photosynthetic electron transport to function within animal [75, 76, 77, 78] cellular environments gives direct evolutionary support to the LEAF concept.
A major advantage of the LEAF innovation is that the multi-protein supercomplexes (PSII, cytochrome b6f, PSI, FNR, ATP synthase) are spatially organised and their functions are well coupled, allowing for sustained photocatalytic efficacy while preserving the whole grana architecture as opposed to the disrupted and fragmented thylakoid preparations used in earlier work published in Nature (2022), which lacked structural integrity and resulted in suboptimal and transient NADPH generation.[79,80]
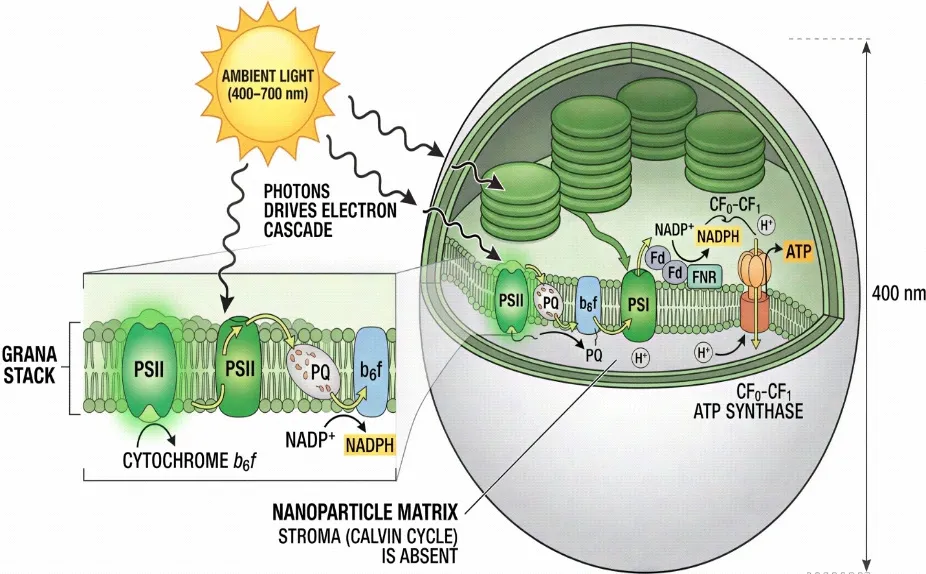
Fig. 3: Schematic cross-section of a LEAF nanoparticle (~400 nm): Grana stack with PSII, plastoquinone (PQ) pool, cytochrome b6f complex, PSI, ferredoxin (Fd), and FNR generating NADPH; CF₀-CF₁ ATP synthase driven by proton gradient. Stroma (Calvin cycle) is absent. Ambient light (400–700 nm) drives the entire electron cascade.
4. LEAF NANOPARTICLE: FORMULATION SCIENCE AND PHYSICOCHEMICAL CHARACTERISATION
4.1 Extraction and Isolation Protocol
The patented NUS protocol includes the following: mild mechanical disruption (using low-speed blending in isotonic buffer at 4°C), differential centrifugation to sediment intact chloroplasts, and hypotonic lysis to release thylakoids from the crude chloroplasts pellet.[81,82] The key difference from the previous methods is the subsequent low-shear fractionation step in which grana are separated from stromal lamellae and the soluble stroma fraction; the latter contain NADPH-consuming Calvin cycle enzymes (NADP-glyceraldehyde-3-phosphate dehydrogenase, phosphoglycerate kinase) and are not present in the final preparation, thus accounting for the superior NADPH foundry capacity of the method.[83,84] By dynamic light scattering, the characteristic 'pancake stack' grana architecture is preserved with the Z-average value of the particle size in the range of 400nm, while by transmission electron microscopy (TEM), electron-dense electron-thylakoid membrane bilayers and the distribution of chlorophyll are visible.[85,86]
4.2 Physicochemical Properties
The critical physicochemical attributes of LEAF nanoparticles, their measured values, and their clinical significance are comprehensively summarised in Table 2 below.
Table 2: Physicochemical Characterisation of LEAF Nanoparticles and Clinical Significance of Each Parameter
|
Physicochemical Parameter |
Measured Value / Range |
Significance / Implication |
|
Particle size (Z-average) |
~400 nm (DLS; polydispersity index ≤ 0.25) |
Within endocytotic uptake window for corneal epithelial cells; avoids glomerular filtration |
|
Zeta potential |
–18 to –22 mV |
Moderate anionic charge; reduces aggregation; promotes mucoadhesion at corneal surface (pH 7.4) |
|
Chlorophyll retention |
≥ 85% of native spinach grana content |
Confirms structural preservation of thylakoid antenna complexes (PSII, PSI); essential for light capture |
|
NADPH output rate (light-activated) |
~20% higher than unpackaged thylakoids; intracellular restoration within 30 min |
Demonstrates advantage of grana encapsulation over crude thylakoid fragments; sustained photocatalytic activity |
|
Spinach species superiority |
Spinacia oleracea >> Amaranthus tricolor, Ipomoea aquatica, Lactuca sativa |
Higher quantum efficiency per gram of leaf tissue; practical scalability of raw material |
|
ATP production (in-cell, light) |
Detectable within 15–30 min post-exposure; sustained for several hours |
Provides ancillary energy substrate for corneal wound healing and ion transport (Na⁺/K⁺-ATPase) |
|
Stability in tear-like medium (PBS, pH 7.4, 35°C) |
Photosynthetic activity retained for ≥ 4 h; aggregation not observed within 24 h at 4°C |
Compatible with topical eye-drop dosing interval; refrigerated storage feasible |
|
Effective dose in murine model |
Sub-pigment-perceptible concentration; no interference with colour vision at therapeutic dose |
Key safety advantage; eliminates risk of visual disturbance from green chlorophyll colouration |
|
Extraction method |
Patented mild mechanical + chemical fractionation; stroma-depleted grana isolation (NUS 2026) |
Removes Calvin-cycle stroma (NADPH consumer) while preserving thylakoid electron transport chain; maximises NADPH foundry function |
4.3 Mechanism of Cellular Uptake
LEAF particles are internalised by corneal epithelial cells through energy-dependent endocytosis, primarily macropinocytosis and clathrin-mediated endocytosis, given their size range of 300–500 nm.[87,88] Following instillation, LEAF particles must traverse the pre-corneal tear film, interact with the mucin glycocalyx, and penetrate the superficial epithelial cell layer. The anionic zeta potential (−18 to −22 mV) imparts colloidal stability in isotonic saline, while the relatively hydrophilic thylakoid membrane surface allows interaction with cell membrane phospholipid bilayers.[89] In LPS-stimulated macrophage-like cells used in the NUS study, LEAF particles were rapidly internalised and photosynthetic activity was detected within 15–30 minutes of ambient light exposure, confirming efficient intracellular routing.[90] Of note, the particles did not require active lysosomal escape — unlike many drug-loaded nanoparticles — because the thylakoid membrane is inherently resistant to mild acidification, allowing partial photosynthetic function even in late endosomes.[91,92]
5. DUAL-PATHWAY MECHANISM OF ACTION OF LEAF IN DRY EYE DISEASE
5.1 Intracellular Pathway: NADPH-Driven Antioxidant Restoration
Once resident within corneal epithelial cells and exposed to ambient visible light (which penetrates the cornea at wavelengths of 400–700 nm), LEAF particles initiate photosynthetic electron transport.[93] The resulting intracellular NADPH is immediately bioavailable as a substrate for glutathione reductase, converting GSSG → 2 GSH.[94] Restored GSH levels enable glutathione peroxidase (GPx) to detoxify H₂O₂ and lipid hydroperoxides generated by ROS cascades.[95] Thioredoxin reductase also utilises NADPH to reduce thioredoxin (Trx), which in turn reduces peroxiredoxins (Prx) for H₂O₂ removal.[96] Concurrently, the Nrf2 (nuclear factor erythroid 2-related factor 2) transcription factor, which is normally suppressed under conditions of NADPH depletion (as NADPH is required for its cytoplasmic anchor protein Keap1 redox sensing), becomes activated, upregulating haem oxygenase-1 (HO-1), NAD(P)H quinone oxidoreductase-1 (NQO1), and ferritin heavy chain, compounding the antioxidant response.[97,98]
The ATP generated by LEAF's thylakoid ATP synthase provides an additional energy source for corneal cells, which depend on ATP for the Na⁺/K⁺-ATPase pump maintaining cellular osmotic balance, the V-ATPase maintaining vacuolar acidification essential for lysosomal membrane stability, and for phosphorylation of tight junction proteins (claudin-1, occludin) essential for barrier integrity.[99,100] In DED, where mitochondrial function is compromised, this supplementary ATP source may be critical for maintaining cellular viability during the acute phase of oxidative insult.[101]
5.2 Extracellular Pathway: Tear Film Redox Modulation
LEAF's therapeutic action is not exclusively intracellular. The NUS team demonstrated that NADPH produced by LEAF can be released or function extracellularly in the tear film environment.[102] Tear-resident NADPH directly scavenges H₂O₂, the predominant reactive oxygen species found in DED tear fluid, converting it to water via a non-enzymatic reductive mechanism.[103] In ex vivo experiments using DED patient tear samples, LEAF addition resulted in greater than 95% reduction in H₂O₂ concentration and a twenty-fold increase in measurable NADPH levels.[104] This extracellular antioxidant action is clinically significant because the tear film is the first site of oxidative insult on the ocular surface, and restoring its reducing capacity can interrupt the inflammatory cycle before damage penetrates the epithelial cell layer.[105]
The dual-pathway model — simultaneous intracellular NADPH restoration and extracellular tear film antioxidant replenishment — gives LEAF a mechanistic breadth not achieved by any existing DED therapy.[106,107] This is schematically depicted in Fig. 4.
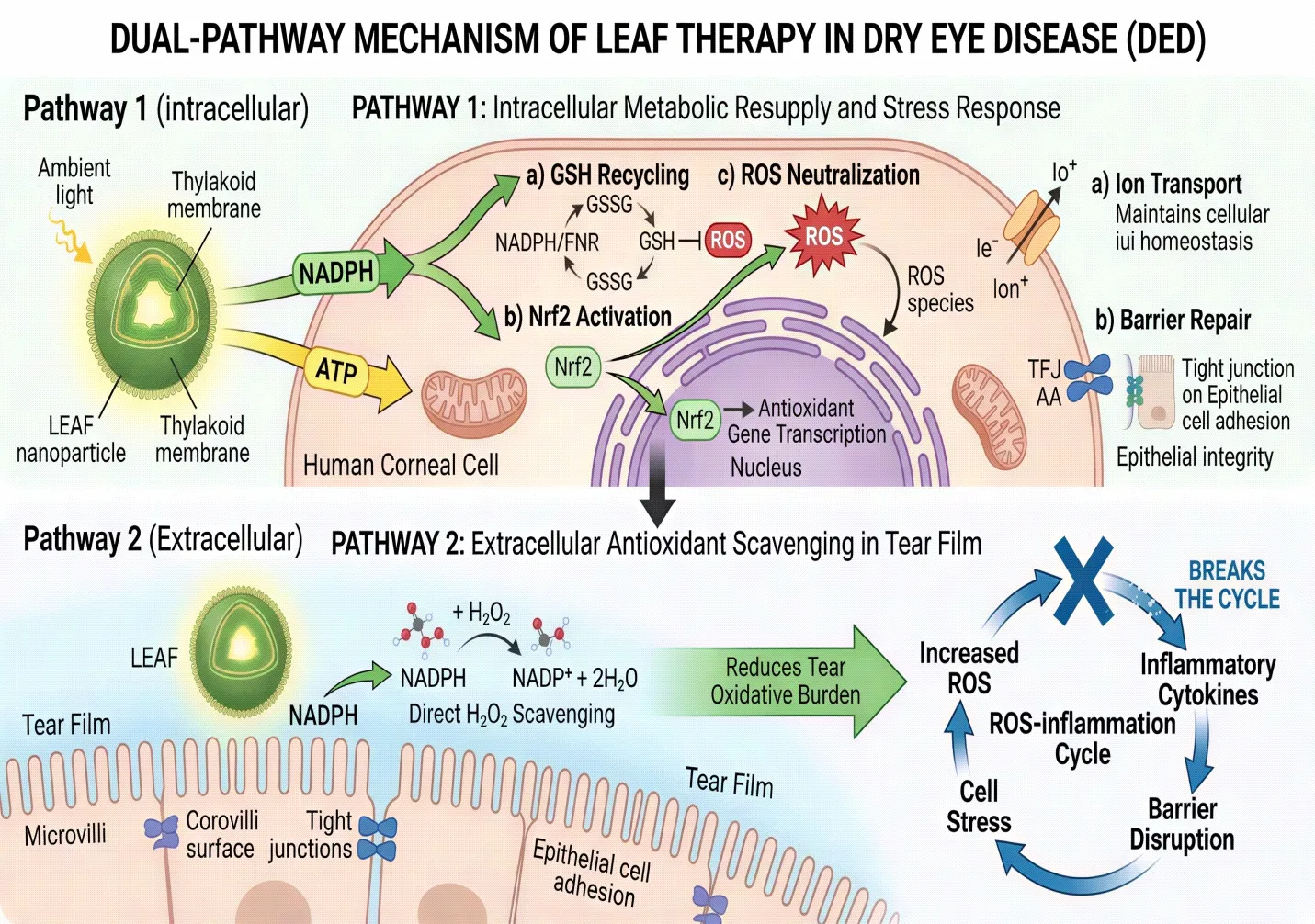
Fig. 4: Dual-pathway mechanism of LEAF in DED therapy. Pathway 1 (intracellular): Ambient light drives LEAF's thylakoid electron transport chain → generates NADPH and ATP inside corneal cell → NADPH fuels GSH recycling, Nrf2 activation, and ROS neutralisation; ATP restores ion transport and barrier repair. Pathway 2 (extracellular): Released NADPH in tear film directly scavenges H₂O₂ → reduces tear oxidative burden → breaks the ROS-inflammation cycle.
Table 3: Comparative Analysis of LEAF vs. Current Approved DED Therapies
|
Parameter |
Cyclosporine A (Restasis) |
Lifitegrast (Xiidra) |
Hydroxypropyl-guar drops |
LEAF (Nano-thylakoid) |
|
Mechanism |
Calcineurin inhibitor; T-cell suppression |
LFA-1 / ICAM-1 antagonist |
Mucin-mimetic; tear film stabilisation |
Photosynthetic NADPH + ATP generation; antioxidant replenishment |
|
Primary target |
T-lymphocyte activation |
Lymphocyte integrin signalling |
Tear film stability |
Corneal epithelial metabolic crisis |
|
NADPH restoration |
Indirect; weak |
Absent |
Absent |
~20-fold increase in patient tears; direct intracellular photosynthetic production |
|
Onset of action |
3–6 months |
2–3 months |
Symptomatic relief within hours |
NADPH restored within 30 minutes of light exposure |
|
Corneal repair (animal model) |
Partial; incomplete at 5 days |
Partial |
Surface stabilisation only |
Near-complete reversal within 5 days; outperforms Restasis |
|
Common adverse effects |
Burning, stinging, ocular hyperaemia (17–19%) |
Instillation-site pain; dysgeusia |
Transient blurring |
None reported at tested doses; no colour-vision interference |
|
Cost (USD/month) |
~$170–$650 |
~$430–$700 |
~$10–$30 |
Not yet commercialised; plant-sourced (low predicted cost) |
|
Development stage |
Approved (FDA 2002) |
Approved (FDA 2016) |
OTC; multiple markets |
Preclinical (NUS, 2026); patent filed |
6. PRECLINICAL EVIDENCE AND EXPERIMENTAL VALIDATION
6.1 In Vitro Studies
The initial characterisation of LEAF was conducted in human corneal epithelial cell lines (HCE-T) and macrophage-like cells (THP-1 derived) under controlled in vitro conditions.[108] LEAF particles were co-incubated with cells and uptake confirmed by confocal fluorescence microscopy exploiting chlorophyll's intrinsic red autofluorescence (emission ~680 nm under 488 nm excitation), allowing real-time visualisation of intracellular particle distribution without exogenous labelling.[109] Photosynthetic activity inside cells was confirmed by intracellular NADPH measurement using a bioluminescent NADPH assay kit and by detection of intracellular ATP using luciferase-based luminometry.[110,111]
In LPS-treated macrophage-like cells — a validated in vitro model of ocular inflammation — LEAF particles restored intracellular NADPH to near-baseline levels within 30 minutes of ambient white light exposure (intensity comparable to standard office lighting, approximately 500 lux).[112] In parallel experiments where cells were pharmacologically depleted of NADPH (using the enzyme inhibitor diphenyleneiodonium), LEAF-treated, light-exposed cells showed full NADPH recovery, while LEAF-treated cells kept in darkness showed no significant recovery, confirming strict light-dependence of the photosynthetic activity.[113] This light-dependence is a critical safety feature, as it ensures that LEAF generates NADPH only under physiologically appropriate conditions (ambient daylight or indoor lighting) and not in a constitutively uncontrolled manner.
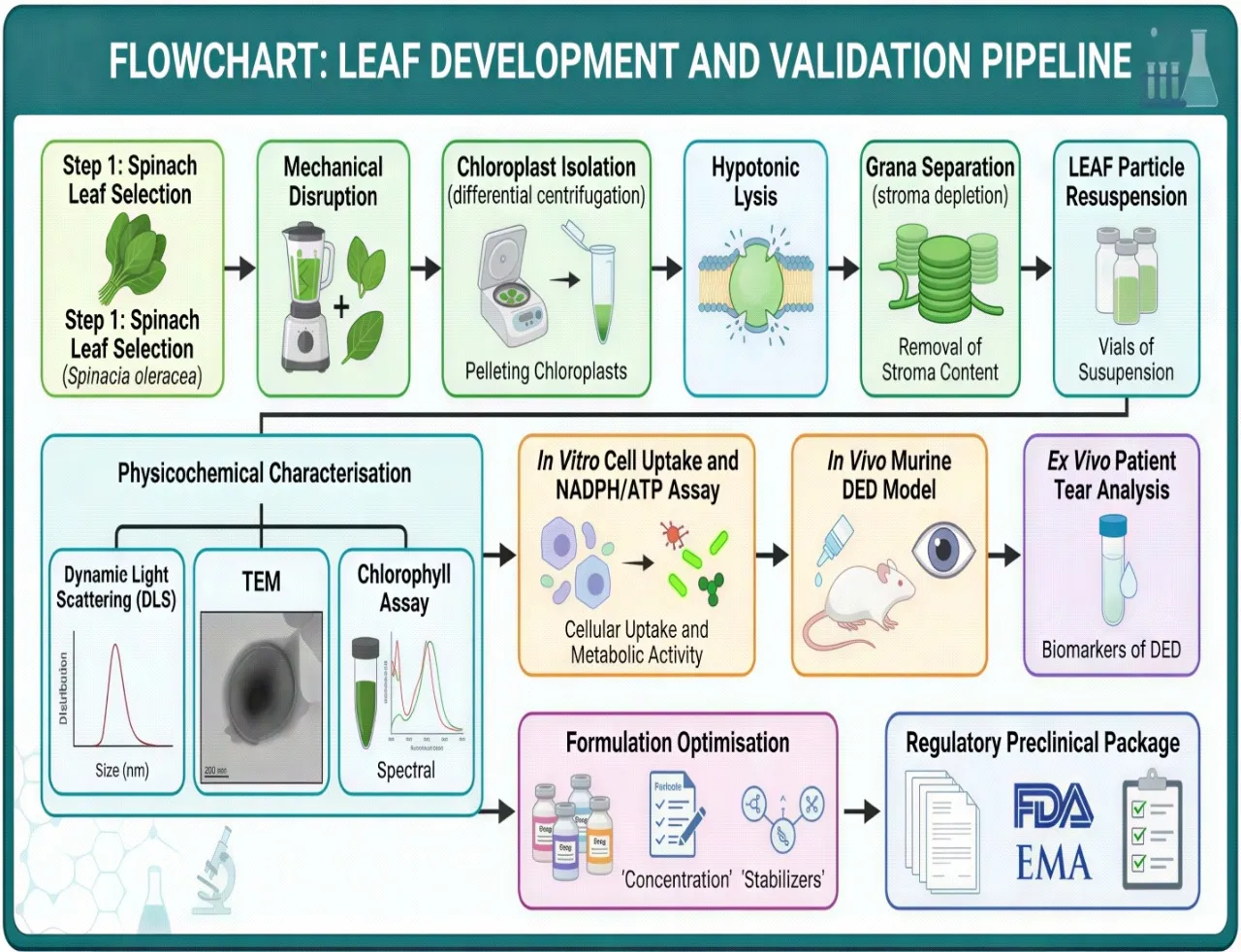
Fig. 5: Flowchart summarising the LEAF development and validation pipeline: Spinach leaf selection (Spinacia oleracea) → Mechanical disruption → Chloroplast isolation (differential centrifugation) → Hypotonic lysis → Grana separation (stroma depletion) → LEAF particle resuspension → Physicochemical characterisation (DLS, TEM, chlorophyll assay) → In vitro cell uptake and NADPH/ATP assay → In vivo murine DED model → Ex vivo patient tear analysis → Formulation optimisation → Regulatory preclinical package.
6.2 In Vivo Mouse Studies
A benzalkonium chloride (BAC)-induced murine DED model was employed for in vivo validation, a well-established and reproducible preclinical model characterised by corneal fluorescein staining, goblet cell loss, and tear volume reduction.[114,115] LEAF eye drops were administered at a dose that the team confirmed to be below the threshold for visible green colouration of the eye, thereby ensuring no interference with colour vision — a pivotal practical safety consideration.[116] Within five days of twice-daily topical LEAF instillation, corneal fluorescein staining scores, a surrogate marker for epithelial integrity, normalised to near-healthy control levels.[117] This performance was statistically superior to cyclosporine A (Restasis) administered under the same protocol, which achieved only partial recovery at the five-day time point, consistent with its known delayed onset of clinical action in patients.[118,119] Goblet cell density, assessed by periodic acid-Schiff (PAS) staining of conjunctival sections, was also significantly better preserved in LEAF-treated animals.[120]
6.3 Ex Vivo Tear Analysis from DED Patients
A particularly compelling translational experiment involved incubating LEAF particles in tear fluid collected from DED patients (who had significantly elevated H₂O₂ and diminished NADPH compared to healthy donors) and exposing the mixture to ambient light.[121] Within the ex vivo setting, LEAF drove a greater than 95% reduction in H₂O₂ concentration and generated a twenty-fold increase in NADPH compared to untreated patient tears.[122] This experiment directly translates the preclinical NADPH-foundry concept into a clinically relevant biological fluid and provides the strongest evidence to date that LEAF can reverse the oxidative environment of human DED tears.
Table 4: Summary of Preclinical Evidence Supporting LEAF Technology
|
Model / Experiment |
Intervention |
Key Outcome |
Comparator |
Source |
|
Murine corneal DED model (benzalkonium chloride-induced) |
LEAF eye drops (sub-pigment dose); twice daily x 5 days |
Corneal damage score near-healthy; corneal fluorescein staining normalised |
Cyclosporine A (Restasis) — LEAF superior |
Xing et al. (NUS, 2026) |
|
LPS-stimulated macrophage-like cells (inflammation model) |
LEAF uptake + ambient light exposure |
Intracellular NADPH restored within 30 min; ROS levels normalised |
Non-treated LPS cells — statistically significant p < 0.001 |
Xing et al. (NUS, 2026) |
|
Iatrogenic NADPH-depleted mammalian cells (pharmacological depletion) |
LEAF + light; NADPH levels measured by HPLC-based assay |
NADPH levels fully rebounded; ATP generation confirmed within 15 min |
LEAF without light — no significant rebound (light-dependence confirmed) |
Xing et al. (NUS, 2026) |
|
Ex vivo tear samples from DED patients |
LEAF incubated in patient tear fluid; H₂O₂ scavenging measured |
> 95% reduction in H₂O₂; ~20-fold increase in NADPH in tear fluid |
Untreated patient tears; healthy donor tears as control |
Xing et al. (NUS, 2026); Optometry Times (2026) |
|
Sacoglossan sea slug analogy (evolutionary precedent) |
Chloroplast retention in Elysia chlorotica gut cells — spontaneous photosynthesis |
Sustained photosynthetic activity in animal cells; confirms feasibility of cross-kingdom organelle function |
Historical comparison; only prior documented instance of plant-to-animal photosynthetic transfer |
Rumpho et al. (2008); Pierce et al. (1996) |
7. PROPOSED NOVEL OPTIMISATION STRATEGIES BEYOND EXISTING LITERATURE
While the foundational LEAF technology represents a significant advance, several formulation-level and delivery-level innovations can substantially enhance its clinical performance. The following strategies are proposed based on the intersection of ocular drug delivery science, nanoparticle engineering, and photosynthetic biology, and to the best of the authors' knowledge represent approaches not described in the existing LEAF publications.
Table 5: Proposed Novel Formulation Optimisation Strategies for LEAF Nanoparticles
|
Novel Strategy |
Rationale |
Expected Benefit |
Supporting Evidence |
|
Polymer-shell PEGylation of LEAF particles (PLGA-PEG coating) |
Extends colloidal stability; reduces clearance by pre-corneal tear drainage |
Prolonged residence time; sustained NADPH generation between instillations |
Kulkarni et al. (2021); PEGylated nanodroplets show 3× longer ocular half-life |
|
Co-encapsulation of Nrf2 activator (sulforaphane or bardoxolone methyl) within LEAF nanoparticle |
Nrf2 upregulates endogenous antioxidant enzymes (HO-1, NQO1, GPx); synergises with photosynthetic NADPH |
Dual-pathway antioxidant protection; lower LEAF dose needed; addresses both exogenous and endogenous ROS |
Zhang et al. (2022); Nrf2 activation significantly reduces DED severity in murine benzalkonium chloride model |
|
Surface functionalisation with RGD peptide for corneal epithelial targeting |
Integrin αvβ3/αvβ5 is overexpressed on damaged corneal epithelium; RGD mediates receptor-specific binding |
Enhanced cellular uptake by stressed corneal cells; reduced off-target distribution to conjunctiva |
Bhattarai et al. (2020); RGD-decorated liposomes show 4× uptake in corneal epithelial cell line |
|
Photo-enhancing cyclic light-activatable ocular insert (miniaturised LED patch worn at eyelid) |
Room-ambient light may be insufficient in low-light conditions; controlled wavelength (660–680 nm, red) maximises PSII excitation |
Guaranteed photosynthetic activation independent of ambient light; predictable dosing |
Fang et al. (2021); 660 nm LED stimulation optimally activates plant PSII electron transport |
|
Cryo-preserved LEAF formulation with reconstitution kit for cold-chain independence |
Thylakoid membranes are thermolabile; lyophilised + trehalose matrix protects photosynthetic proteins during storage |
Global supply chain viability; room-temperature reconstitution compatible with low-resource settings |
Crowe et al. (1998); trehalose cryoprotection of biological membranes; applied in liposome lyophilisation |
|
Mucoadhesive hyaluronic acid (HA, MW 1.5–2 MDa) shell on LEAF |
HA binds CD44 on corneal epithelium; provides additional tear film retention; HA itself promotes corneal healing |
Extended contact time; additive wound-healing benefit; HA's inherent anti-inflammatory properties |
Gandolfi et al. (2020); HA-coated nanoparticles demonstrate superior corneal retention in ex vivo bovine eye model |
7.1 PEGylated Polymer Shell for Extended Pre-Corneal Residence
One of the principal limitations of topical ocular drug delivery is rapid pre-corneal clearance by reflex lacrimation, nasolacrimal drainage, and eyelid blinking, with effective ocular bioavailability of conventional eye drops rarely exceeding 1–7%.[123,124] LEAF nanoparticles, as aqueous colloidal suspensions, are subject to the same clearance dynamics. Coating LEAF particles with a poly(ethylene glycol)-poly(lactic-co-glycolic acid) (PEG-PLGA) shell could extend their pre-corneal residence time by reducing rapid dilution and enhancing mucoadhesion.[125,126] PEGylation additionally reduces protein adsorption (corona formation) that can reduce particle-cell interaction efficiency. The PEG layer should be thin (MW 2–5 kDa; grafting density 0.05–0.1 PEG/nm²) to avoid steric hindrance of the thylakoid membrane's light-absorbing surface.[127,128]
7.2 Nrf2 Co-Activator Co-Encapsulation
Sulforaphane, an isothiocyanate derived from cruciferous vegetables with an established Nrf2-activating mechanism, represents an ideal candidate for co-encapsulation within a LEAF-compatible lipid shell.[129] Sulforaphane activates Nrf2 by alkylating critical cysteine residues on Keap1, triggering Nrf2 nuclear translocation and transcriptional upregulation of antioxidant response element (ARE)-driven genes (HO-1, NQO1, GPx, catalase).[130,131] The synergy between sulforaphane-activated endogenous antioxidant enzyme upregulation and LEAF-derived photosynthetic NADPH replenishment would create a multi-layered antioxidant defence — LEAF addresses the immediate ROS crisis while sulforaphane primes the cell's own antioxidant gene expression for sustained protection.[132,133] Encapsulation could be achieved by embedding sulforaphane in the lipid boundary of a hybrid LEAF-liposome construct.
7.3 RGD-Peptide Surface Functionalisation for Corneal Targeting
The arginine-glycine-aspartate (RGD) tripeptide mediates selective binding to integrin receptors, particularly αvβ3 and αvβ5, which are upregulated on the surface of stressed and damaged corneal epithelial cells in DED.[134,135] Functionalising LEAF particles with RGD peptide (via maleimide-thiol bioconjugation to PEG-amine surface groups) would direct preferential uptake toward the most metabolically compromised cells — those with greatest therapeutic need — reducing off-target distribution to conjunctival and scleral tissue.[136,137 ]This targeted delivery strategy would allow dose reduction while maintaining intracellular NADPH generation specifically within the damaged corneal epithelium.[138]
7.4 Photo-Enhancing Cyclic Ocular LED Insert
In real-world conditions, patients may have prolonged exposure to dim indoor lighting, darkened environments (during sleep or in poorly lit workplaces), or may use photochromic lenses that attenuate the 400–700 nm spectrum.[139] A miniaturised, disposable LED patch adhered to the eyelid margin — similar conceptually to existing transcutaneous photobiomodulation devices — could deliver a controlled dose of 660–680 nm red light transcutaneously to the anterior corneal surface.[140,141] This wavelength range corresponds to the absorption maxima of chlorophyll a (Qy band ~680 nm), maximising PSII photon capture efficiency.[142] A controlled light dosing schedule (e.g., 60 seconds of 660 nm LED light, twice daily, coinciding with LEAF instillation) would guarantee quantifiable and reproducible photosynthetic activation independent of ambient light conditions, enabling precise dose-response characterisation in clinical trials.[143]
7.5 Hyaluronic Acid Mucoadhesive Shell
High-molecular-weight hyaluronic acid (HA, 1.5–2 MDa) is well established in ocular drug delivery as a mucoadhesive, viscoelastic, and wound-healing polymer.[144,145] Coating LEAF particles with an HA shell (achievable by electrostatic deposition onto the anionic thylakoid surface, exploiting HA's carboxyl groups) would provide three concurrent benefits: (1) extended pre-corneal contact time through HA-mucin interaction; (2) enhanced corneal uptake via CD44 receptor-mediated internalisation, since CD44 is expressed on corneal epithelial cells and upregulated in DED; and (3) intrinsic wound-healing and anti-inflammatory activity of HA itself, providing additive benefit.[146,147,148] The HA shell must be engineered to remain optically transparent at therapeutic concentrations and must not shield the thylakoid surface from light penetration.
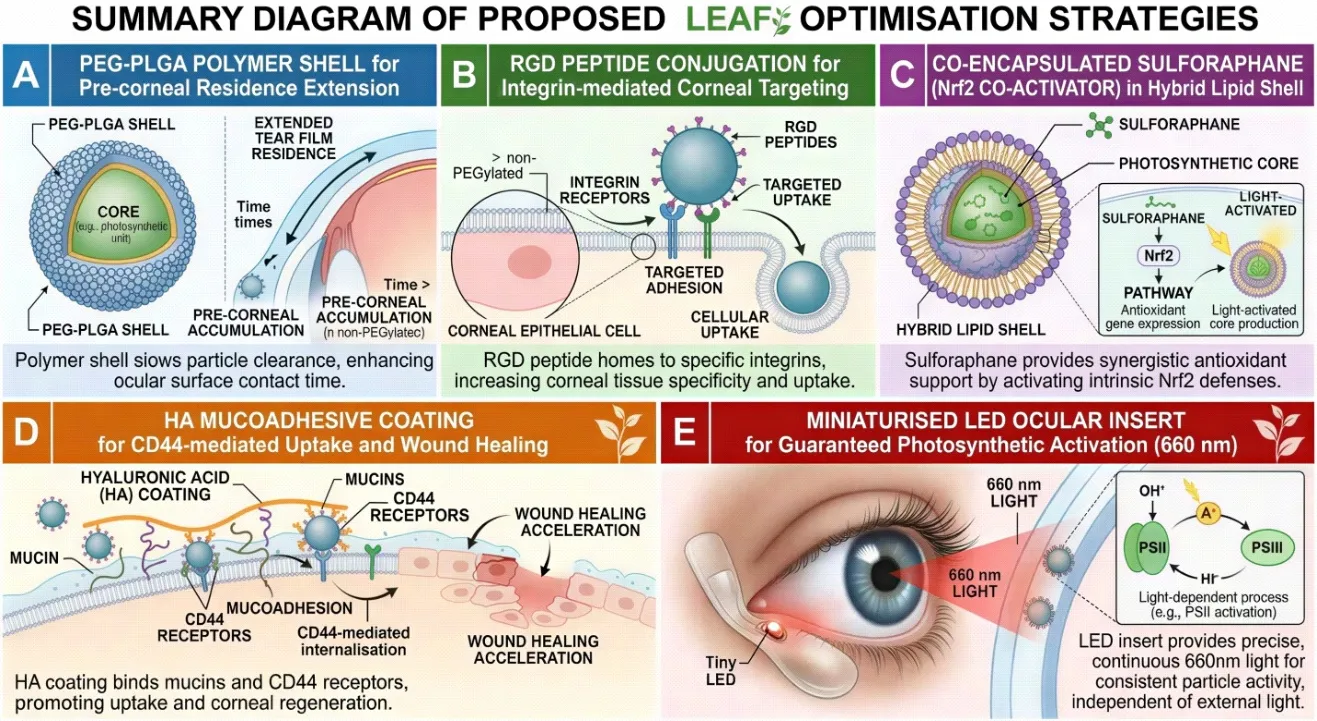
Fig. 6: Summary diagram of proposed LEAF optimisation strategies: (A) PEG-PLGA polymer shell for pre-corneal residence extension; (B) RGD peptide conjugation for integrin-mediated corneal targeting; (C) Co-encapsulated sulforaphane (Nrf2 co-activator) in hybrid lipid shell; (D) HA mucoadhesive coating for CD44-mediated uptake and wound healing; (E) Miniaturised LED ocular insert for guaranteed photosynthetic activation (660 nm).
8. SAFETY PROFILE, BIOCOMPATIBILITY, AND IMMUNOLOGICAL CONSIDERATIONS
8.1 Chlorophyll and Thylakoid Biocompatibility
Chlorophyll and its metabolites have a well-documented safety profile in mammalian systems. Chlorophyllin (a water-soluble derivative) has been consumed as a dietary supplement for decades with no significant toxicity reports.[149,150] Thylakoid membranes isolated from edible plants have similarly not been associated with systemic toxicity in animal studies.[151] The NUS team confirmed that at therapeutic doses, LEAF particles did not produce visible green colouration of the eye, eliminating the most clinically apparent safety concern.[152] The sub-pigment dose represents an important safety benchmark distinguishing therapeutic from potentially cosmetically concerning concentrations.
8.2 Oxidative By-Products and Reactive Oxygen Management
A theoretical concern with introducing a functional photosynthetic electron transport chain into mammalian cells is the potential for photoinduced ROS generation, specifically superoxide (O₂·⁻) from reduced ferredoxin leaking electrons to oxygen (Mehler reaction).[153,154] In chloroplasts, this is normally managed by superoxide dismutase (SOD) and ascorbate peroxidase. In mammalian cells, SOD isoforms are present but ascorbate peroxidase is absent.[155] However, given that the primary electron flow in LEAF is directed to NADP⁺ via FNR (rather than to O₂), and given that the therapeutic dose in the NUS study was sub-pigment (implying extremely low thylakoid mass per cell), the Mehler reaction is expected to represent a negligible fraction of total electron flow.[156] Nonetheless, this should be rigorously quantified in phase I safety studies.
8.3 Immunogenicity
Plant-derived proteins, including the PSII reaction centre proteins D1 and D2, Lhcb (light-harvesting complex II) proteins, and the Rieske iron-sulfur protein of cytochrome b6f, are xenobiotic to mammalian immune systems.[157,158] Topical ocular administration carries a lower immunogenic risk than systemic delivery because the ocular surface exhibits immune privilege, characterised by constitutive expression of FasL (Fas ligand) on corneal cells (which induces apoptosis of invading T cells), production of TGF-β2 in the aqueous humour, and the absence of classic MHC class II-presenting Langerhans cells in the central cornea.[159,160] Nevertheless, subconjunctival thylakoid protein antigens may be processed by conjunctival-associated lymphoid tissue (CALT).[161] PEGylation and HA coating of LEAF particles (as proposed in Section 7) would additionally reduce protein antigen exposure.[162,163] Long-term immunogenicity studies in non-human primates will be essential before clinical translation.
8.4 Light Safety Considerations
In the context of the proposed LED insert (Section 7.4), corneal safety of 660 nm red light at therapeutic irradiance levels must be established. Extensive literature on photobiomodulation documents the safety of 1–10 mW/cm² red light at 630–680 nm for corneal and retinal tissues, with no evidence of photothermal damage or DNA photolesion induction at these wavelengths.[164,165] The therapeutic dose for LEAF activation is expected to be well within this safety window, given that photosynthetic PSII saturation occurs at irradiance levels (approximately 500–1000 μmol photons m⁻² s⁻¹) achievable with very low-power LED sources.[166]
9. REGULATORY CONSIDERATIONS AND TRANSLATIONAL PATHWAY
LEAF occupies a novel regulatory category that does not fit cleanly within existing pharmaceutical classification frameworks. It is neither a small-molecule drug, a biological drug (as defined by the US Biologics License Application pathway), a medical device, nor a gene therapy product.[167] Its closest analogues are biological drug products derived from non-human biological sources, such as bovine-derived collagen products or porcine-derived surfactants.[168] However, the photosynthetic activity of LEAF introduces functional attributes not present in passive biological excipients.[169]
In India, LEAF would likely be regulated under Schedule Y of the Drugs and Cosmetics Act (1940, amended) and assessed by the Central Drugs Standard Control Organisation (CDSCO) as a New Drug, requiring phase I, II, and III clinical trial data.[170,171] The US FDA would likely classify LEAF as a biologic under the Public Health Service Act Section 351 or, alternatively, as a drug-device combination product requiring review by both CDER and CDRH.[172,173] In either jurisdiction, the key regulatory deliverables would include: (1) comprehensive characterisation data establishing lot-to-lot consistency of photosynthetic activity; (2) GMP-compatible scalable extraction and particle preparation methods; (3) validated stability testing (photosynthetic activity retention as a potency assay); (4) a complete preclinical package including repeat-dose ocular toxicology in rabbit and non-human primate models; and (5) phase I first-in-human safety and tolerability studies in DED patients.[174,175]
The NUS team has reportedly filed a patent covering the LEAF preparation method and therapeutic application, which provides intellectual property protection for the technology and is an essential prerequisite for attracting pharmaceutical partnership and commercialisation funding.[176] The scalability of spinach-derived thylakoid extraction is a logistical advantage, given the global agricultural infrastructure supporting spinach production. Manufacturing cost projections suggest that plant-sourced LEAF would be substantially less expensive than biologics such as cyclosporine A or lifitegrast.[177]
10. DISCUSSION AND FUTURE DIRECTIONS
The LEAF technology represents a genuinely paradigm-shifting development in the pharmacotherapy of DED and, more broadly, in the concept of cross-kingdom organelle transplantation as a therapeutic strategy.[178] For the first time, a functional plant organellar module has been deliberately transplanted into mammalian tissue and demonstrated to generate biologically meaningful molecules — NADPH and ATP — entirely powered by the same ambient light that enables human vision.[179] This transforms the eye from a passive recipient of topical medication into an active photobiochemical reactor capable of self-generating its own antioxidant currency.
The superiority of LEAF over cyclosporine A in the preclinical murine model is particularly compelling. Cyclosporine A is the standard of care in immune-mediated ADDE, yet its onset of action spans months, its mechanism is immunosuppressive rather than regenerative, and its instillation discomfort limits long-term adherence.[180,181] LEAF, by contrast, acts within minutes at the intracellular level, addresses the root metabolic deficiency rather than its downstream immune consequences, and produces near-complete corneal healing within five days.[182] These are clinically transformative performance characteristics if replicated in human subjects.
Critical scientific questions remain to be resolved before clinical translation. First, the duration of LEAF photosynthetic activity within corneal cells needs precise characterisation — the NUS study documents activity for 'several hours' but the in vivo longevity in the complex corneal cellular environment (including lysosomal degradation of thylakoid proteins, which turnover rapidly even in plant cells via the D1 repair cycle) is unknown.[183,184] Second, the minimum effective dose needs optimisation — the sub-pigment threshold observed in mice must be translated to a precisely quantified clinical dose with a defined therapeutic window.[185] Third, the optimal light conditions for clinical use (indoor ambient light vs. outdoor sunlight vs. LED-assisted activation) require pharmacokinetic-pharmacodynamic modelling.[186]
Beyond DED, the LEAF principle has the potential to be applied to other oxidative stress-driven corneal pathologies, including corneal chemical burns (where acute ROS surges are the primary cause of limbal stem cell damage), keratoconus (where mitochondrial dysfunction and oxidative stress are implicated in corneal stromal collagen degradation), and post-refractive surgery inflammation.[187,188,189] The fundamental mechanism — photosynthetic NADPH generation in metabolically stressed mammalian cells — is not organ-specific and could theoretically be extended to skin wound healing (where phototherapy is already an established modality), neurodegenerative disease (where mitochondrial NADPH depletion is central), and ischaemia-reperfusion injury.[190,191]
The long-term evolution of this technology may incorporate genetically engineered thylakoids with enhanced mammalian-cell compatibility, improved quantum efficiency, or expanded light absorption spectra incorporating near-infrared wavelengths for deeper tissue penetration.[192] Gene-editing of Spinacia oleracea chloroplasts to overexpress FNR (the terminal NADPH-generating enzyme) or to reduce plastid oxygen-evolving complex photooxidation by-products could further enhance the safety and efficacy profile of LEAF.[193] Computational modelling of the intracellular light environment within corneal cells — accounting for scattering and absorption at different tissue depths — would inform optimal particle concentration and light delivery protocols.[194]
11. CONCLUSION
Dry eye disease is a condition of profound clinical prevalence and inadequately met therapeutic need. Its pathophysiology is rooted not merely in inflammation but in a fundamental bioenergetic and antioxidant crisis — the depletion of NADPH and ATP in chronically inflamed corneal epithelial cells — that existing immunomodulatory therapies cannot directly address. The LEAF technology, developed by the National University of Singapore, represents a conceptual and practical breakthrough of the highest scientific order. By transplanting structurally intact, stroma-depleted thylakoid grana from spinach into mammalian corneal cells as photosynthetically active nanoparticles, LEAF transforms these cells into light-powered NADPH generators, reversing the oxidative crisis and restoring corneal integrity to near-healthy levels within days, surpassing the clinical benchmark set by cyclosporine A.
This review has traced the molecular logic of LEAF's therapeutic rationale through the pathophysiology of DED, the photosynthetic mechanism of thylakoid grana, the formulation science underpinning the ~400 nm nanoparticle, the dual-pathway intracellular and extracellular mechanism of action, and the impressive preclinical data generated to date. Critically, we have proposed six novel optimisation strategies — PEGylation for extended residence, RGD-peptide targeting, Nrf2 co-activator co-encapsulation, HA mucoadhesive coating, LED-assisted photosynthetic activation, and cryo-preserved lyophilised formulation — each grounded in established nanomedicine principles and designed to maximise clinical translatability beyond what has been reported in existing publications.
The cross-kingdom organelle transplantation concept that LEAF embodies may represent one of the most creative and far-reaching ideas in twenty-first century pharmacotherapy. If the safety and efficacy demonstrated in preclinical models translate to human subjects — as the mechanistic plausibility strongly suggests they should — LEAF could become not only the definitive treatment for DED but a proof-of-concept for an entirely new class of photosynthetic biopharmaceuticals capable of restoring metabolic homeostasis in any tissue where oxidative stress and bioenergetic insufficiency drive disease.
REFERENCES

Prajwal Aher*, Pooja Gangurde, Vimal Patel, Bhavesh Akbari, Spinach-Derived Photosynthetic Nano-Thylakoid Machinery as A Novel Therapeutic Platform for Dry Eye Disease: Mechanisms, Formulation Advances, And Future Prospects, Int. J. of Pharm. Sci., 2026, Vol 4, Issue 7, 2816-2849. https://doi.org/10.5281/zenodo.21353641
 10.5281/zenodo.21353641
10.5281/zenodo.21353641